


1Shanxi Key Laboratory of Stem Cells for Immunological Dermatosis, Institute of Dermatology, Taiyuan Central Hospital of Shanxi Medical University, Taiyuan, Shanxi, and 2Dermatology Hospital, Southern Medical University, Guangdong, China
Although it is known that psoriatic dermal-derived mesenchymal stem cells (DMSCs) dysregulate keratinocyte proliferation, the biological activity profile of keratinocytes influenced by psoriatic DMSCs remain unknown. In the present study, we assessed the impact of psoriatic DMSCs on keratinocyte proliferation, differentiation, and glucose metabolism in normal human epidermal keratinocytes co-cultured with or without psoriatic DMSCs. Co-culture of normal human epidermal keratinocytes with psoriatic DMSCs down-regulated expression levels of proteins associated with cell junction assembly (alpha-actinin-1, catenin beta-1, poliovirus receptor-related protein 4 and procollagen-lysine, 2-oxoglutarate 5-dioxygenase 2), while upregulating proteins associated with keratinocyte proliferation and differentiation (involucrin, isoform 2 of Histone-binding protein, isoform 3 of Telomeric repeat-binding factor 2 and keratin 13). Moreover, co-culture of normal human epidermal keratinocytes with psoriatic DMSCs stimulated keratinocyte proliferation and glycolysis, but reduced keratinocyte junctions. Taken together, these results demonstrate that psoriatic DMSCs increase keratinocyte proliferation and glycolysis, and reduce cell junctions, suggesting a pathogenic role of psoriatic DMSCs in epidermal hyperplasia, aberrant differentiation, and reduction in turnover time of keratinocytes in psoriasis.
Key words: psoriasis; dermal mesenchymal stem cells; keratinocytes; cell junctions; cell proliferation; glycolysis.
Accepted Apr 2, 2020; Epub ahead of print Apr 8, 2020
Acta Derm Venereol 2020; 100: adv00122.
Corr: Kaiming Zhang, Shanxi Key Laboratory of Stem Cells for Immunological Dermatosis, Taiyuan Central Hospital of Shanxi Medical University, No.5, east third lane, Jiefang road, Taiyuan, Shanxi 030009, China. E-mail: zhangkaiming@sina.com
Psoriasis is a chronic inflammatory skin disease that affects approximately 2% of the world’s population. The promotion of keratinocyte proliferation by psoriatic dermal-derived mesenchymal stem cells is abnormal. However, the biological activity profile of keratinocyte influenced by dermal-derived mesenchymal stem cells remain unclear. In this study, the differentially expressed proteins were enriched in cellular glucose homeostasis, tricarboxylic acid metabolic process, cell junction assembly, keratinocyte proliferation and differentiation. Moreover, increased keratinocyte proliferation, increased glycolysis and reduced cell junc-tions were observed with psoriasis treatment. Our study suggests that psoriatic dermal-derived mesenchymal stem cells induce keratinocyte proliferation and glycolysis, and reduce cell junctions.
Psoriasis is a chronic inflammatory skin disease, affecting approximately 2% of the world’s population (1). Dysregulations in the proliferation and differentiation of keratinocytes (KCs) are the key characteristics of psoriasis (2, 3). For a long time, the pathogenesis of psoriasis has been a hotspot for dermatological research. However, the underlying mechanism is still unclear.
Mesenchymal stem cells (MSCs) are capable of multi-directional differentiation and self-renewal, and increasingly attract research attention. The functions of MSCs include, but are not limited to, promoting wound healing (4), inhibiting T-cell proliferation (5), affecting angio-genesis (6) and inducing KC proliferation (7). These biological effects are mainly mediated by direct cell-to-cell contact and, in particular, paracrine effects (8). Dermal mesenchymal stem cells (DMSCs) are the main MSCs in the dermis. Our previous studies demonstrated that DMSCs from psoriatic patients displayed multiple abnormalities, including mRNA expression, gene methy-lation (9, 10), patterns of secreted cytokines (11) and their inhibitory effects on lymphocytes (12). Moreover, MSCs from psoriatic lesions secreted high concentrations of nitric oxide (13). Together, these results suggest that DMSCs play a critical role in the pathogenesis of psoriasis, and are directly involved in various biological activities in the local microenvironment. Recent studies have shown that MSCs can differentiate into keratinocytes (14), suggesting that MSCs can dysregulate KCs in psoriasis. This assumption is further supported by the observation that co-cultures of psoriatic DMSCs and HaCaT cells stimulate the proliferation of HaCaT cells (15).
To further investigate the effect of DMSCs on KC activity, here we measured the protein levels, cell junctions, cell proliferation and energy metabolism in normal human epidermal keratinocytes (NHEKs) co-cultured with DMSCs. Additionally, to avoid tumour genic effects, we replaced the immortalized HaCaT cell line (used in previous studies) with NHEKs, derived from normal primary KCs.
Skin samples
Twelve Chinese Han patients with quiescent psoriasis vulgaris with plaque lesions who were admitted to undergo plastic surgery at Taiyuan City Centre Hospital (5 women and 7 men, mean age 37.4 years), and 12 healthy controls (6 women and 6 men, mean age 39.5 years) were enrolled in this study. Diagnosis of psoriasis was based on both clinical and pathological features. Psoriasis Area and Severity Index scores (PASI) were between 5.8 and 13.8 (mean 8.63). All patients were with no other autoimmune diseases and relevant family history, and had not been on treatment for at least 6 months prior to skin sample collection. Volunteers exhibited no systemic or autoimmune diseases. All of samples (12 psoriasis and 12 controls) were about 1 × 0.5 cm, surgically taken from the chest or the back. Skin samples from psoriatic lesions were with erythema and scale. Ethical approval was obtained from the Medical Ethics Committee of Taiyuan City Centre Hospital. Work was carried out according to Declaration of Helsinki, and all subjects provided written informed consent.
Isolation and expansion of dermal-derived mesenchymal stem cells
The DMSCs were isolated from both healthy volunteers and psoriatic patients, followed by expansion and identification, as described previously (12). Briefly, the dermis was separated from the epidermis by incubation of skin samples in 0.25% dispase (Sigma-Aldrich, St Louis, MO) for 2–4 h at 4°C, and the cells were incubated at a concentration of 1.0×105 cells/ml in DMEM/F12 (1: 1; HyClone Laboratories, Losan, UT) containing 5% foetal bovine serum (FBS, HyClone), 10 ng/ml basic fibroblast growth factor (bFGF, Sigma), 10 μl/ml B27, and 1% antibiotic (penicillin/streptomycin (10,000 U/ml), Gibco) at 37°C in a humidified atmosphere supplemented with 5% CO2. Non-adherent cells were removed after 72 h, and half of the medium was changed every 3–4 days. Adherent DMSCs were passaged at 80% confluence. The purity of the DMSCs was verified by cell surface markers using a flow cytometer (Beckman Coulter, Inc., CA) (negative for CD34, CD45, HLA-DR, but > 95% positive for CD73, CD90, CD44, CD29, CD105, data not shown) at passage 3 (P3).
Co-culture of dermal-derived mesenchymal stem cells with normal human epidermal keratinocytes
Total of 1×105 adult pooled NHEK cells (Promo Cell, Heidelberg, Germany) were cultured in keratinocyte serum-free medium (Promo Cell) with epidermal growth factor plus bovine pituitary extract (Gibco, Gran Island, NY) and 1% antibiotics, and seeded at 1×104 cells/ml in 25 cm2 tissue culture flasks at 37°C in 5% CO2. The medium was changed daily until the cells were confluent. NHEKs were detached using 0.25% trypsin and 0.04% EDTA. Total of 3×105 NHEKs (2×105 cells/ml) were seeded onto Transwell plates at passages 2–4 to co-culture with 1×105 DMSCs from 12 psoriasis and 12 control (500μl cell suspension at a concentration of 2×105 cells/ml), respectively. NHEKs were detached using 0.25% trypsin-0.04% EDTA after 72 h of co-culture. Afterward, NHEKs were harvested for further analyses. NHEKs cultured without any DMSCs for 72h alone served as “Untreated NHEKs”. The NHEKs co-cultured with psoriatic DMSCs and control DMSCs were labelled as “P-DMSC-Treated NHEKs” and “C-DMSC-Treated NHEKs”, respectively.
Quantitative analysis of protein expression using iTRAQ
In addition to untreated NHEKs, NHEKs co-cultured with 5 psoriatic DMSCs (labelled “P-DMSCs-Treated NHEKs”) and 5 control DMSCs (“C-DMSCs-Treated NHEKs”) were pooled for iTRAQ. Protein was extracted, using an ultrasonic disruptor with pyrolysis liquid. Following centrifugation, the supernatant was treated sequentially with 10 mM DTT, 55 mM IAM, 100 μg trypsin/sample, and then the iTRAQ tag. After mixing, tagged peptides were separated and gradient eluted, using an SCX column. The entire eluant was processed at 214 nm absorbance, and twelve components were obtained following filtering. Each component was desalinated, using a StrataX desalination column and dried. Dried components were separately analysed, using Triple TOF 5600 LC-ESI-MSMS. Five microliters of each component (approximately 2.5 μg protein) was separated by liquid chromatography. For data analysis, a reference database was established, using peak lists of standards. And peptides and proteins in each sample were subsequently identified. After comparing relative expression levels between samples by t-test, key proteins of interest were identified.
Simple Western analysis
Simple Western analysis were performed on a Wes system according to the ProteinSimple introduction using a 12-230 kDa Separation Module (ProteinSimple, Silicon Valley, USA) and the Anti-Rabbit Detection Module (ProteinSimple). Briefly, cell lysate samples were mixed with a master mix and heated at 95°C for 5 min. The samples, blocking reagent, primary antibodies (Abcam, Cambridge, UK), secondary antibodies, chemiluminescent substrate, and separation and stacking matrices were also dispensed to designated wells in a 25-well plate. Instrument default settings were used: stacking and separation at 475 V for 30 min; blocking reagent for 5 min, primary and secondary antibody both for 30 min; Luminol/peroxide chemiluminescence detection for 15 min (exposures of 1-2-4-8-16-32-64-128-512s). The digital image was analysed with Compass software (ProteinSimple). The expression of β-actin was analysed as an internal reference for relative expression.
Fluorescence redistribution after photobleaching
FRAP was used to detect cell junctions between adjacent KCs. Total of 1×106 NHEKs from 3 conditions were seeded into confocal culture dishes. After growing to about 80% confluence, the cells were washed with 0.01 M PBS, and incubated with 6-carboxyfluorescein diacetate (10 µg/ml, Sigma-Aldrich) for 10 min at 37°C, followed by wash and suspension in 100 μl DMEM/F12. FRAP was detected with a confocal microscope (Olympus, Japan). The dye was excited at 488 nm, and the emission spectrum was recorded at 570 nm. Before bleaching, the target cells were selected and scanned with a lower intensity laser for 3 frames. The selected cells were then bleached by exposure to 50 scans at a laser intensity of 95%, and the recovery of fluorescence in the bleached cells was measured every 15s for a total of 5 min. After correction for background bleaching, the recovery of fluorescence in the bleached cells at 5 min was compared with that of the prebleached scans to calculate the recovery percentage.
Cell Count Kit-8 assay
First, 100 µl of cell suspension were seeded into 96-well plates at 1×105 cells/ml. After culture for 24 h at 37°C, 10 µl of CCK8 solution (Wuhan Boster Biological Technology., Wuhan, China) was added to each well, followed by incubation for 1 h. The optical density (OD) at 450 nm was measured with a microplate reader, and analysed to reflect cell proliferation.
The measurements of oxygen-consumption rate and extracellular acidification rate
Cellular oxidative phosphorylation and glycolysis were detected by Mito Cell Stress, and glycolysis Stress was assessed with Seahorse XF Cell Metabolic Analyzers (Seahorse Bioscience, Inc.). Briefly, 1 × 106 cells were seeded in 24-well plates matched for XF24. Before measurements, the cells were washed and incubated with unbuffered media in a CO2-free incubator for 24 h. The detection of OCR was performed in XF Base Media after addition of following drugs to the cultures: oligomycin (1.0 μM), carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP) (2.0 μM) and rotenone/antimycin A (1 μM). The detection of extracellular acidification rate (ECAR) was performed in XF Base Media, and the following drugs were added: glucose (1.0 μM), oligomycin (1.0 μM) and 2-deoxyglucose (0.5 μM). The oligomycin-sensitive oxygen-consumption rate (OCR) (OxPhos OCR), an indicator of mitochondria ATP production, was the difference of OCR prior to, and after addition of oligomycin. Likewise, the glycolytic ECAR, a product of glycolysis, was the difference of ECAR prior to, and after addition of 2-deoxyglucose (2-DG).
Statistical analysis
Except for iTRAQ, all of the data were analysed first by the F-test for variance homogeneity and then compared using a paired Student’s t-test. Analyses were performed using SPSS 17.0. A value of p < 0.05 was considered statistically significant.
Protein expression
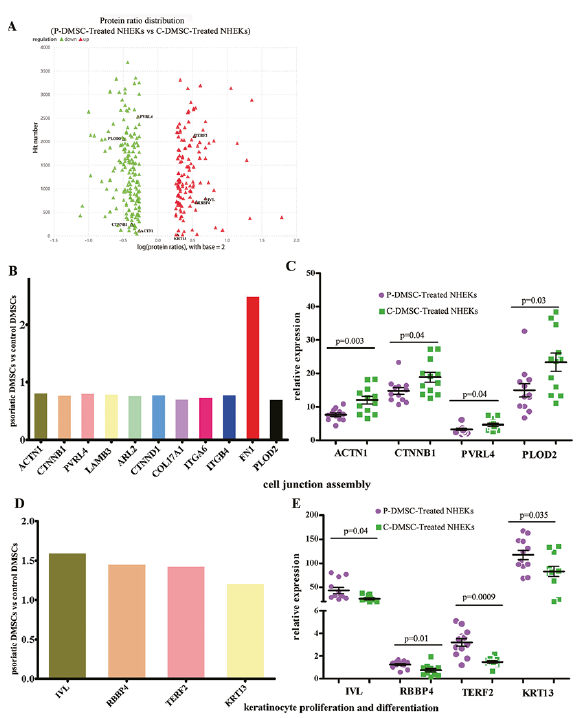
We obtained 406,980 signals in the mass spectrum experiment. After analysis using Mascot software, we identified 49,874 matched pieces, including 42,718 copies of the unique spectra. In total, 4,431 proteins and 21,533 peptides were identified, including 19,653 unique peptides. In comparison to C-DMSC-Treated NHEKs, with the threshold of fold changes ≥ 1.2 and p < 0.05, 297 differentially expressed proteins were identified in P-DMSC-Treated NHEKs (Fig. 1A, Table S1). These proteins were enriched for the tricarboxylic acid cycle (p < 0.00001), cellular glucose homeostasis (p = 0.002), cell junction assembly (p = 0.006), keratinocyte differentiation (p = 0.003911912) and keratinocyte proliferation (p = 0.026). Notably, 9 of 10 differentially expressed proteins associated with cell junction assembly were downregulated in P-DMSC-Treated compared to C-DMSC-Treated NHEKs (Fig. 1B, C, Fig. S1A), including alpha-actinin-1 (ACTN1, fold change 0.83), catenin beta-1 (CTNNB1, 0.79), poliovirus receptor-related protein 4 (PVRL4, 0.82), laminin subunit beta-3 (LAMB3, 0.80), ADP-ribosylation factor-like protein 2 (ARL2, 0.78), catenin delta-1 (CTNND1, 0.80), collagen alpha-1 (XVII) (COL17A1, 0.72), isoform Alpha-6X1A of Integrin alpha-6 (ITGA6, 0.75) and integrin beta (ITGB4, 0.80). Additionally, procollagen-lysine, 2-oxoglutarate 5-dioxygenase 2 (PLOD2), which is critical for the stability of intermolecular collagen crosslinks (GeneCards 2019), was also downregulated following the treatment with psoriatic DMSCs (fold change 0.71). Moreover, in comparison to control DMSCs, co-culture of NHEKs with psoriatic DMSCs markedly lowered expression levels of proteins associated with keratinocyte proliferation and differentiation, including involucrin (IVL, fold change 1.61), isoform 2 of histone-binding protein (RBBP4, 1.47), isoform 3 of telomeric repeat-binding factor 2 (TERF2, 1.44), and keratin 13 (KRT13, 1.22) (Fig. 1D, E, Fig. S1B).
To validate the above results, we examined 8 proteins by Simple Western analysis. The results showed that psoriatic DMSCs treatment downregulated expression levels of ACTN1 (7.61 vs 12.00, p = 0.003), CTNNB1 (14.81 vs 18.83, p = 0.04), PVRL4 (3.27 vs 4.61, p = 0.04), and PLOD2 (15.03 vs 23.33, p = 0.03), while increasing expression levels of IVL (40.14 vs 25.61, p = 0.04), RBBP4 (1.22 vs 0.80, p = 0.01), TERF2 (3.26 vs 1.57, p = 0.0009), and KRT13 (116.80 vs 88.34, p = 0.035), consistent with the iTRAQ results.
Fig. 1. Protein expression of iTRAQ and the Simple Western Analysis. A. Protein ratio distribution (P-DMSC-Treated NHEKs vs. C-DMSC-Treated NHEKs); B. Expression of proteins associated with cell junction assembly assessed by iTRAQ (t-test, data are expressed as mean); C. Expression of proteins with associated cell junction assembly assessed by Simple Western Analysis (t-test, data are expressed as mean with standard error of mean (SEM)), D. Expression of proteins associated with keratinocyte proliferation and differentiation assessed by iTRAQ (t-test, data are expressed as mean), E. Expression of proteins associated with keratinocyte proliferation and differentiation assessed by Simple Western Analysis (t-test, data are expressed as mean with SEM).
Psoriatic dermal-derived mesenchymal stem cells decrease cell junctions
To determine the functions of these differentially expressed proteins, next we examined the cell junctions, proliferation and glucose metabolism. As shown in Fig. 2, adjacent cells displayed a higher fluorescence recovery rate (FRR) in comparison to isolated cells in all of 3 conditions (Untreated NHEKs: 16.01% in adjacent cells vs. 5.08% in isolated cells; C-DMSC-Treated NHEKs: 19.13% in adjacent cells vs. 8.32% in isolated cells; P-DMSC-Treated NHEKs: 7.21% in adjacent cells vs. 3.39% in isolated cells; p < 0.05 for all), indicating the presence of cell junctions between the adjacent cells. Co-culture of NHEKs with psoriatic DMSCs significantly decreased the FRR of the bleached cells (7.21% vs 16.01%, p < 0.05), while FRR of the bleached cells was comparable in NHEKs co-cultured with or without control DMSCs (19.13% vs 16.01%, p > 0.05). Moreover, the FRR of NHEKs treated with psoriatic DMSCs was significantly lower than that treated with control DMSCs (7.21% vs 19.13%, p < 0.05). The FRAP results suggest an inhibitory effect of psoriatic DMSCs on KC junctions.
Fig. 2. Psoriatic dermal-derived mesenchymal stem cells (DMSCs) decreased intercellular junction between adjacent keratinocytes (KCs) based on 1×106 cells. (A) Recovery after fluorescence bleaching within 5 min in Untreated normal human epidermal keratinocytes (NHEKs). (B) Recovery after fluorescence bleaching within 5 min in P-DMSC-treated NHEKs. (C) Recovery after fluorescence bleaching within 5 min in C-DMSC-treated NHEKs. (D) Fluorescence redistribution after photobleaching in 3 conditions ((Untreated NHEKs, P-DMSC-treated NHEKs and C-DMSC-treated NHEKs). t-test, data are expressed as Mean ± standard deviation). Note: The red arrow indicates the bleached cells.
Psoriatic dermal-derived mesenchymal stem cells stimu-late keratinocyte proliferation
Keratinocyte proliferation was detected by CCK8 assays (Fig. 3A). Co-culture of NHEKs with psoriatic DMSCs for 24 h significantly increased OD (450 nm) values (0.67 vs 0.45 in Untreated NHEKs, and vs. 0.54 in C-DMSC-Treated NHEKs, p < 0.05 for both). In contrast, co-culture of NHEKs with control DMSCs only induced a minimal increase in OD values (0.54 vs 0.45, p > 0.05). These results demonstrate that psoriatic DMSCs stimulate KC proliferation.
Psoriatic dermal-derived mesenchymal stem cells alter glucose metabolism in normal human epidermal keratinocytes
Because psoriasis is often complicated with metabolic syndrome, we assessed next whether psoriatic DMSCs could alter glucose metabolism in NHEKs. Indeed, co-culture of NHEKs with psoriatic DMSCs induced more dramatic increase in ECAR, an indicator of glycolysis, although control DMSCs also caused a moderate increase (Fig. 3C). Likewise, psoriatic DMSCs significantly increased glycolysis and glycolytic reserve capacity (Fig. 3C). However, co-culture of NHEKs with either psoriatic or control DMSCs increased OCR (Fig. 3B). These results indicate that psoriatic DMSCs alter glucose metabolism in keratinocytes.
Fig. 3. The cell proliferation and metabolic readiness of normal human epidermal keratinocytes (NHEKs) in 3 conditions (Untreated NHEKs, P-DMSC-treated NHEKs and C-DMSC-treated NHEKs). (A) The OD (450 nm) of NHEKs by a CCK8 assay based on 1×104 cells (t-test, data are expressed as mean with standard deviation (SD)). (B) The oxygen-consumption rate (OCR) based on 1×106 NHEKs detected at different time points in three conditions (Data are expressed as mean with SD). OCR under oligomycin, carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP), and antimycin A/rotenone treatments (Data are expressed as Mean with SEM). (C) The extracellular acidification rates (ECAR) based on 1×106 NHEKs detected at different time points in 3 conditions (Data are expressed as mean with SD). ECAR under glucose, oligomycin, and 2-DG.
Psoriasis, a recurrent skin disease that seriously affects patients’ physical and mental health, is characterized by a dysregulation of KC proliferation (2) and differentiation (3), along with a shortened epidermal turnover time (16). Recently, role of skin-derived MSCs in the pathogenesis of psoriasis has attracted much attention. Prior studies showed that injection of human umbilical cord blood-derived MSCs alleviated psoriasis-like skin inflammation in mice (17). Orciani et al. (13) reported that VEGF levels were highest in medium conditioned by psoriatic perilesional MSCs, while increased nitric oxide concentration was observed in medium conditioned by MSCs isolated from psoriatic lesion. We showed here that psoriatic DMSCs decreased expression levels of cell junction assembly in NHEKs, including ACTN1, CTNNB1, PVRL4 and PLOD2, which all play important role in keratinocyte function. ACTN1 is critical for cell junctions, such as myosin-II-driven contractility, formation of cell-cell and cell-ECM adhesion complexes, and tight junctions (18–20). CTNNB1, an integral protein of adherens junctions, also plays a role in cell-cell adhesion by regulating cadherin-mediated cell adhesion and the interplay of adherens junction proteins with the actin cytoskeleton (21), while PVRL4 is a cell adhesion molecule promoting the formation of adherens junctions through the nectin-afadin and cadherin-catenin complexes (22). Moreover, PLOD2 catalyses the hydroxylation of lysyl residues in collagen-like peptides, a process required for the stability of intermolecular collagen crosslinks (GeneCards 2019). Therefore, psoriatic DMSC-induced dysregulation of proteins associated with cell junction assembly likely contributes to reductions in KC junction in psoriasis, leading to alteration in KC differentiation (3) and shortened epidermal turnover time (16).
One of the main characters of psoriasis is KC hyperproliferation, which can be induced by pro-inflammatory cytokines, such as TNF-α, IL-1β, IL-6 (23). In addition, MSCs can also stimulate KC proliferation and migration, in part, via upregulation of IL-8 expression in dermal fibroblasts (7, 24). In the present study, we demonstrated that psoriatic DMSCs upregulated at least 4 cell proliferation- and differentiation-related proteins, including IVL, RBBP4, TERF2 and KRT13. Among these proteins, RBBP4 binds directly to retinoblastoma protein, regulating cell proliferation and possibly being involved in transcriptional repression of E2F-responsive genes (25). TERF2 is present in telomeres at the metaphase of the cell cycle, regulating telomere length and the protective activity of telomeres (26). Thus, psoriatic DMSC-induced upregulation of IVL, RBBP4, TERF2 and KRT13 expression could also contribute to dysregulations of KC hyperproliferation and differentiation.
The present study also showed that psoriatic DMSCs increased expression levels of proteins involved in the progression of the tricarboxylic acid cycle and cellular glucose homeostasis. Glycolysis is a key metabolic pathway that provides energy for cellular activities and consumes equivalents to sustain cell division (27). Glycolysis also plays an essential role in cell proliferation by maintaining high levels of glycolytic intermediates to support cellular anabolic reactions (28). Conversely, the decreased glycolysis is related to cell differentiation (29, 30), particularly keratinocyte differentiation (27). Hence, increased KC proliferation in psoriasis could be attributable to DMSC-induced elevations in glycolysis.
Despite the capacity of MSCs to differentiate, increasing evidence has shown that much of the disease-modulating activity of MSCs is due to products secreted by the cells. Our previous studies showed that secretion of EGF, SCF, IL-11, bFGF, IL-3, IL-8 and HGF was abnormal in psoriatic DMSCs (11). In the present study, the co-culture method was adapted to avoid direct contact between NHEKs and DMSCs. Thus, the influence of DMSCs on KC junctions, proliferation, differentiation and glucose metabolism is via the paracrine effects, including EGF, SCF, IL-11, bFGF, IL-3, IL-8 and HGF. Moreover, in the present study, we used psoriatic DMSCs at passage 3, which display comparable phenotypes of primary psoriatic DMSCs (31). In addition, the present study showed that control and psoriatic DMSCs differentially regulates keratinocyte function. Therefore, the impact of psoriatic DMSCs on keratinocyte function unlikely represent non-specific effects of DMSCs, but psoriasis-associated specific effects.
In conclusion, psoriatic DMSCs abnormally regulate KC junctions, proliferation, differentiation and glucose metabolism, leading to epidermal hyperplasia, aberrant differentiation and reduced epidermal cell turnover time, which are characteristics of psoriasis (Fig. 4). However, the involvement of cytokines and other molecules in these processes remains to be determined.
Fig. 4. Proposed mechanism of dermal-derived mesenchymal stem cells (DMSCs) participation in psoriasis. Psoriatic DMSCs secrete many cytokines (such as EGF, SCF, IL-11, bFGF, IL-3, IL-8 and HGF), and these cytokines primarily have 4 effects on keratinocye (KC): induction of KC proliferation, induction of KC glycolysis, dysregulation of KC differentiation and reduction of KC junctions. Increased KC glycolysis also induces KC proliferation and reduces KC differentiation. Meanwhile, the decreased KC junctions also dysregulated KC differentiation. Finally, the disequilibrium of KC activity leads to psoriasis characterized by epidermal hyperplasia and aberrant differentiation (32).
This project was supported by the National Natural Science Foundation of China (grant nos. 81773336, 81602768, 81401360 and 81472888). The funding was attribute to the collection, analysis, and interpretation of data.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize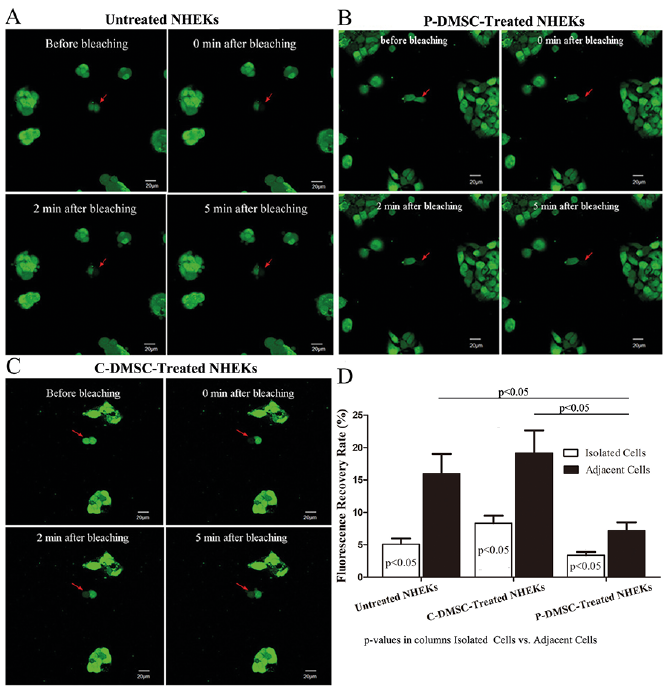 Click to show fullsize
Click to show fullsize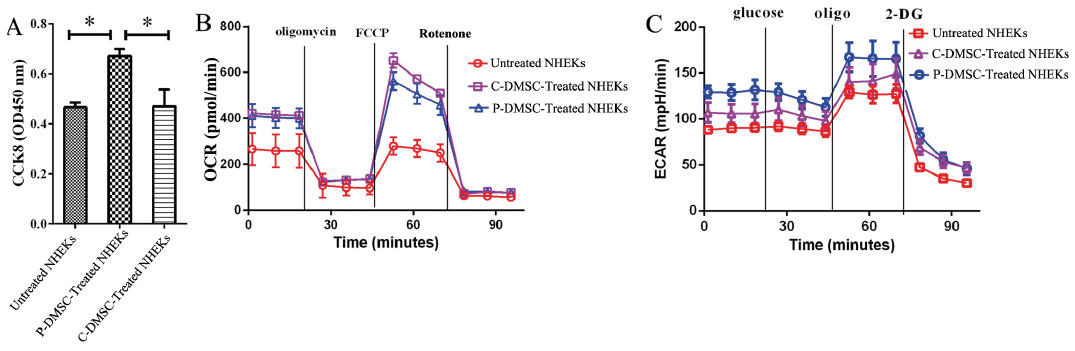 Click to show fullsize
Click to show fullsize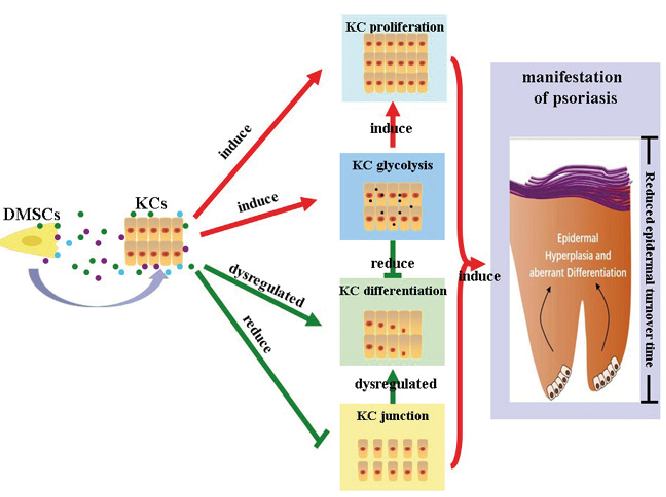 Click to show fullsize
Click to show fullsize