


1University of Veterinary Medicine Hannover, Institute for Pharmacology, Toxicology and Pharmacy, 2University of Veterinary Medicine Hannover, Institute of Anatomy, 3Hannover Medical School, Division of Immunodermatology and Allergy Research, Department of Dermatology and Allergy, Hannover, and 4Institute of Pharmacology and Toxicology, Department of Veterinary Medicine, Freie Universität Berlin, Berlin, Germany
Psoriasis is a chronic inflammatory skin disorder characterized by hyperproliferative keratinocytes and immune cell infiltration into the skin, often accompanied by itch. Histamine, acting via histamine 1–4 receptors, is known to modulate immune responses in the skin and to induce itch. The aim of this study was to test the role of histamine 2 receptors and histamine 4 receptors in the imiquimod-induced psoriasis-like skin inflammation model. BALB/c mice were treated intraperitoneally with amthamine (histamine 2 receptor agonist), JNJ-39758979 (histamine 4 receptor antagonist), a combination of both, or vehicle twice daily in a preventive manner. Imiquimod was applied once daily onto the back skin for 10 consecutive days. Stimulation of histamine 2 receptors and blockade of histamine 4 receptors ameliorated imiquimod-induced skin inflammation. The combination of amthamine and JNJ-39758979 reduced skin inflammation even more, diminished epidermal hyperproliferation, and inhibited spontaneous scratching behaviour. A combination of histamine 2 receptor agonist and histamine 4 receptor antagonists could represent a new strategy for the treatment of psoriasis.
Key words: histamine; psoriasis; itch; H2 receptor; H4 receptor.
Accepted Oct 21, 2020; Epub ahead of print Oct 26, 2020
Acta Derm Venereol 2020; 100: adv00342.
doi: 10.2340/00015555-3674
Corr: Wolfgang Bäumer, Institute of Pharmacology and Toxicology, Department of Veterinary Medicine, Freie Universität Berlin, DE-14195 Berlin, Germany. E-mail: Wolfgang.baeumer@fu-berlin.de
Psoriasis is characterized by hyperproliferative keratinocytes and immune cell infiltration. Histamine has pro-inflammatory properties; it stimulates the proliferation of keratinocytes, and the histamine concentration in skin is increased in psoriatic lesions. However, classical antihistamines (histamine 1 receptor antagonists) play only a minor role in therapy for psoriasis. This study reveals that treatment with histamine 2 receptor agonists, and blockade of the histamine 4 receptor ameliorates inflammation in a mouse model of psoriasis and diminishes psoriasis-associated itch. Histamine 2 receptor agonists and histamine 4 receptor antagonists might thus be new targets to treat psoriasis.
Psoriasis is a chronic autoimmune disease characterized by a hyperproliferative epidermis consisting of premature keratinocytes, incomplete cornification and an elevated immune cell infiltration (1). Psoriasis is mediated by complex interactions between immune cells, especially T cells, antigen presenting cells, and keratinocytes. Histamine might play a pathogenetic role in psoriasis, since histamine increases the proliferation of keratinocytes, and elevated histamine levels can be found in psoriatic skin (2–4). Histamine acts via 4 G-protein-coupled receptors, histamine 1–4 receptors (H1–4R). Whilst activation of H1R and H4R, in general, elicit a pro-inflammatory immune response, activation of H2R is associated with regulatory immune responses. In a mouse model of atopic dermatitis, dual treatment with an H4R and H1R antagonist diminished the inflammatory response in the skin (5). It has been shown that H4R, but not H1R or H2R, modulates the proliferation of keratinocytes (4). Furthermore, plasmacytoid dendritic cells (pDC) isolated from skin and blood of patients with psoriasis highly express H4R. H4R was shown to influence cytokine production and migration of pDC (6). This subpopulation of dendritic cells can produce massive amounts of pro-inflammatory cytokines and are absent in healthy skin (7). According to these findings, H4R antagonists might provide a new strategy to treat psoriasis. However, a recent study by Kim et al. (8) showed that the dual H2R/H4R agonist 4-methylhistamine reduced imiquimod (IMQ)-induced psoriasis-like skin inflammation in mice, arguing against this hypothesis. Thus, the aim of this study was to test the role of H2R and H4R in a mouse model of psoriasis, the IMQ model.
Mice
Female BALB/c mice (BALB/cAnNCrl, 7 ± 1 weeks-old, body weight 20 ± 1 g) were obtained from Charles River (Sulzfeld, Germany). H4R knockout (H4R–/–) mice were generated by Lexicon Genetics (Woodlands Park, TX, USA) and provided by Janssen Research & Development, LLC, La Jolla, CA, USA and backcrossed for more than 10 generations onto the BALB/c strain. The animal experiment has been approved by the Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit, Oldenburg, Germany (AZ. G33.12-42502-04-15/1899). All animals were healthy and were housed in groups of 7 mice per Eurostandard type cages (macrolone, 825 cm² of space), at 22°C with a 12-h light/dark cycle and wood shavings as bedding material. A mouse house and paper tissues as nesting material were used as environmental enrichment. Water and a standard diet (Altromin, Lage, Germany) were available ad libitum.
Imiquimod-induced skin inflammation
Mice were acclimatized to their environment for 2 weeks before the experiments. The animals were randomly allocated into treatment and vehicle group (n = 7 per group). In the experiment with H4R–/– mice, n = 6 H4R–/– mice and n = 6 wild-type mice were used. Sample size calculation was performed by power analysis using G*power 3.1.9.2. for Windows (alpha value 0.5, power 80%, Cohen’s d effect size 0.8). A co-worker blinded to the experimental protocol randomized animals into these groups. Mice were treated with Aldara™ (MEDA Pharma GmbH & Co. KG, Bad Homburg, Germany) cream (containing 5% IMQ) to induce psoriasis-like skin inflammation. This is referred as “IMQ” throughout the current paper. Mice were killed 24 h after the last application of IMQ or soft cream. Mice were killed by inhalation of isoflurane, followed by cardiac puncture for blood collection. Animal experiments were performed in accordance with the recommendations in the ARRIVE guidelines (https://arriveguidelines.org/).
Treatment groups
Five independent experiments were performed. Experiment 1: soft cream vs IMQ (this experiment is referred as the “control” throughout this paper). Soft cream was used as the vehicle cream for the IMQ-containing Aldara™ cream. Experiment 2: Treatment with H2R agonist amthamine administered intraperitoneally (i.p.) and IMQ topically (H2R + IMQ) vs treatment with vehicle (aqua ad injectionem) i.p. and IMQ topically (vehicle + IMQ). Experiment 3: Treatment with H4R antagonist JNJ-39758979 i.p. and IMQ topically (H4R + IMQ) vs treatment with vehicle (aqua ad injectionem) i.p. and IMQ topically (vehicle + IMQ). Experiment 4: Treatment with amthamine plus JNJ-39758979 i.p. and IMQ topically (H2R + H4R+ IMQ) vs treatment with vehicle (aqua ad injectionem) i.p. and IMQ topically (vehicle + IMQ). Experiment 5: Wild-type mice treated with IMQ topically vs H4R–/– mice treated with IMQ topically.
Drug administration
IMQ or soft cream were applied once daily onto the shaved back skin (30 mg). The IMQ model was modified to 30 mg on back skin, since 50 mg induced quite severe inflammation that could be reduced only moderately, even by a strong topical corticosteroid, such as diflorasone-diacetate. H2R agonist amthamine, H4R antagonist JNJ-39758979, a combination of amthamine+JNJ-39758979 or vehicle (aqua ad injectionem) i.p. twice daily for 17 days. Drug administration commenced one week prior to application of IMQ cream. All substances were given at a dosage of 20 mg/kg body weight twice daily.
Assessment of severity of dermatitis
Severity of inflammation of the back skin was evaluated using a modified Psoriasis Area and Severity Index (PASI) by blinded investigators. Mice were assessed individually regarding erythema, scaling and thickening, on a scale from 0 (no symptoms) to 4 (marked symptoms). These 3 individual assessments were summed in a cumulative score from 0 to 12 (total skin score).
Analyses of spontaneous scratching behaviour
Scratching behaviour was determined from day 3 to 10 before topical application of IMQ (i.e. 23 h after the last topical application). Mice were recorded on video for 30 min and scratching bouts on the back and the treated ear with the hind paws were analysed.
Histological analysis of the skin
Lesional skin sites of the back were separated, fixed and paraffin sections stained with haematoxylin and eosin (H&E). Sections were analysed under light microscopy by a blinded observer. Epidermal thickness of 10 randomly selected areas was assessed using Axiovision 4.8 software (Carl Zeiss, Oberkochen, Germany). Total inflammatory cell influx was determined semiquantitatively by a score system ranging from “–” (none) to “’++++”4 (very high cell influx).
Cytokine determination in the skin
Skin was homogenized at first and the total protein concentration of each skin specimen was determined to normalize the protein content between multiple samples. In brief, skin samples were homogenized under liquid nitrogen, transferred to RPMI 1640 medium and 1 mmol/l Pefabloc, and mixed intensively. After centrifugation the supernatants were collected and the protein content was determined with a protein assay. Interleukins (IL)-17A, IL-23, IL-33 and tumour necrosis factor alpha (TNF-α) were measured by enzyme-linked immunoassay (ELISA) according to the manufacturer’s instructions.
Single cell preparation of lymph nodes
The skin-draining lymph nodes (Lnn. axilliares) were isolated and single cell suspensions were counted in a Neubauer chamber.
Data analysis
Five different experiments were performed. Statistical significances between 2 groups were assessed by Mann–Whitney U test using GraphPad Prism version 7.00 for Windows (GraphPad Software, La Jolla, CA, USA). p-values < 0.05 were considered statistically significant. The PASI score and the scratching behaviour were analysed with 2-way analysis of variance (ANOVA) followed by Sidak’s multiple comparisons test. Data given in the text and in the graphs are presented as mean ± standard deviation of the mean (SD). All experiments were conducted/analysed in a blinded manner (all researchers were unaware of which treatment each subject has received until after the experiment was over).
Materials
IMQ (Aldara™ 5% Crème) was obtained from Meda Pharm GmbH & Co. KG (Bad Homburg v.d. Höhe, Germany). As vehicle cream a soft cream was used containing (w/w) of 32.5% paraffin, 22.7% white petrolatum, 3.2% glycerol monostearate 40–50, 2.1%, sorbitanmonostearate, 4.4% polyoxyl ethylene glycol(30)-stearat, 0.1% citric acid monohydrate, 21% H2O, 5% urea, 8.5% glycerin 85% and 0.5% phenoxetol. The vehicle cream was made by a local pharmacy (Apotheke Großer Hillen, Hannover, Germany). Amthamine dihydrobromide was obtained from Tocris Bioscience (Bristol, UK). JNJ-39758979 was provided by Janssen Research & Development (La Jolla, CA, USA). Bovine serum albumin was purchased from Sigma-Aldrich (Darmstadt, Germany). RPMI 1640 medium was purchased from Biochrom GmbH (Berlin, Germany). Pefabloc was obtained from Roche Diagnostics (Basel, Switzerland). The protein assay was obtained from Bio-Rad Laboratories GmbH (München, Germany). All ELISAs (IL-17A, IL-23, IL-33 and f-α) were obtained from R&D Systems (Minneapolis, MN, USA).
Blockade of histamine 4 receptors and stimulation of histamine 2 receptors reduced imiquimod-induced skin inflammation
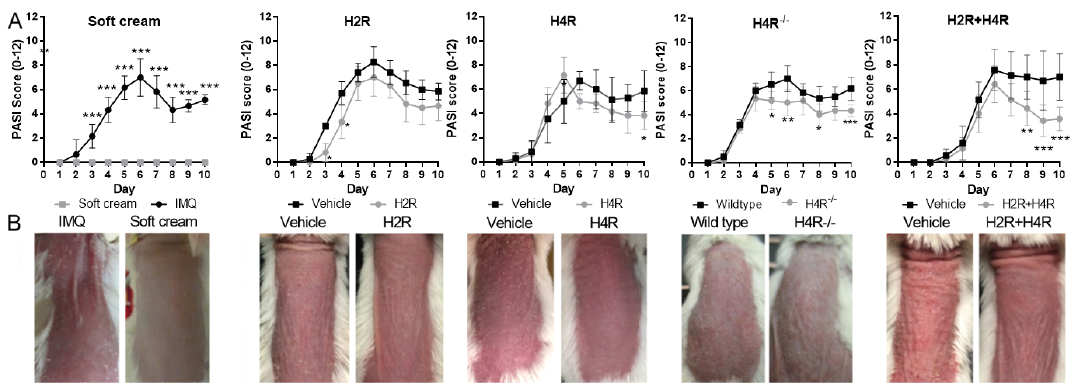
Topical application of IMQ induced local inflammation with erythema, thickening and scaling of the skin (Fig. 1 and Fig. S1). Clinical parameters increased continuously over the first 6 days of application and reached a plateau or decreased slightly by day 7 (Fig. 1). Mice treated with soft cream (vehicle cream) did not develop any visible signs of skin inflammation, and were scored with 0 at any day and any parameter (Fig. 1 and Fig. S1). The total skin score was slightly reduced in mice treated systemically with the H2R agonist amthamine, but only significantly on days 3 and 4. Starting at day 6 of IMQ treatment, the H4R antagonist JNJ-39758979 reduced the total skin score (significantly only on day 10) (Fig. 1A). H4R–/– mice had a lower total skin score starting from day 5 compared with wild-type mice (Fig. 1). The combination of amthamine and JNJ-39758979 led to the strongest reduction in total skin score (Fig. 1). On day 3 after starting the experiment, one of the mice treated with amthamine died immediately after intraperitoneal injection. Necropsy showed that a large blood vessel was inadvertently damaged by the needle.
Fig. 1. Histamine 2 receptor (H2R) stimulation and histamine 4 receptor (H4R) blockade reduced imiquimod (IMQ)-induced skin inflammation. (A) Severity of inflammation of the back skin was evaluated using a modified Psoriasis Area and Severity Index (PASI). Scores (0–4) of the 3 parameters (scaling, thickening, erythema) were summed in a cumulative score from 0 to 12 (total skin score). In mice treated with the vehicle cream (soft cream) instead of IMQ, no signs of skin inflammation were visible. Total skin score was slightly reduced in mice treated with amthamine administered intraperitoneally (i.p.) (H2R) or with JNJ-39758979 i.p. (H4R) compared with the corresponding vehicle groups. Total skin score was reduced in H4R–/– mice compared with wild-type mice. In mice treated with the combination of amthamine and JNJ-39758979 (H2R+H4R) the total skin score was clearly reduced; n = 6 BALB/c mice treated with amthamine and n = 7 BALB/c mice all other treatment groups; n = 6 wild-type and n = 6 H4R–/– mice; *p < 0.05, **p < 0.01, ***p < 0.001. Two-way analysis of variance (ANOVA) followed by Sidak’s multiple comparisons test. Data are presented as mean ± standard deviation (SD). All drugs were given at a dosage of 20 mg/kg i.p. twice daily and were compared with treatment with vehicle (aqua ad injectionem, twice daily i.p.). (B) Clinical signs on day 10 of one representative mouse per group. Mice show erythema, scaling and thickening of the back skin. Mice treated with vehicle cream (soft cream) instead of IMQ did not develop signs of skin inflammation. Clinical signs were less severe in mice treated with the H2R agonist amthamine (H2R), with the H4R antagonist JNJ-39758979 (H4R). H4R–/– mice showed less severe scaling than wild-type mice on day 10. A combination of both (H2R+H4R) compared with the corresponding vehicle groups on day 10 displayed the strongest reduction in lesions.
Combination of an H2R agonist and an H4R antagonist completely abolished scratching behaviour
Spontaneous scratching behaviour was analysed daily (starting at day 3) 23 h after the last application of IMQ. Mice treated with IMQ showed more scratching bouts compared with mice treated with soft cream (Fig. 2A). In amthamine + IMQ-treated mice, the scratching response was not significantly altered compared with vehicle + IMQ-treated mice (Fig. 2B). In mice treated with JNJ-39758979 + IMQ the scratching response was reduced only slightly, if at all (Fig. 2C). The combination of amthamine and JNJ-39758979 almost completely abolished the IMQ-induced scratching behaviour (Fig. 2D). Comparable to the lack of efficacy of the H4R antagonist alone, also in H4R–/– mice, no difference in the scratching behaviour was detected compared with wild-type mice (Fig. 2E).
Fig. 2. Combined treatment with amthamine and JNJ-39758979 administered intraperitoneally (i.p.) inhibited imiquimod (IMQ)-induced scratching behaviour. (A) Mice treated with IMQ showed an increased scratching behaviour compared with mice treated with soft cream. (B) Amthamine i.p. (H2R+IMQ) did not reduce the IMQ-induced scratching behaviour. (C) In mice treated with JNJ-39758979 i.p. + IMQ topically (H4R+IMQ), the scratching response was lower than in mice treated with vehicle i.p. + IMQ topically, but only on days 3 and 6, the scratching behaviour was significantly reduced. (D) The combination of amthamine and JNJ-39758979 i.p. (H2R+H4R+IMQ) almost completely inhibited IMQ-induced scratching behaviour compared with the corresponding vehicle group. (E) IMQ-induced scratching response did not differ between wild-type and H4R–/– mice. **p < 0.01, ***p < 0.001 2-way analysis of variance (ANOVA) followed by Sidak’s multiple comparisons test. n = 6 BALB/c mice treated with amthamine and n = 7 BALB/c mice all other treatment groups; n = 6 wild-type and n = 6 H4R–/– mice. Data are presented as mean ± standard deviation of the mean (SD). All drugs were given in a dosage of 20 mg/kg i.p. twice daily and were compared with treatment with vehicle (aqua ad injectionem, twice daily i.p.).
Epidermal thickening was lower in mice treated with H2R agonist or H4R antagonist
Amthamine and JNJ-39758979-attenuated IMQ-induced epidermal thickening. IMQ strongly increased the epidermal thickness, from approximately 20 ± 3 µm in mice treated with soft cream (control) to 60 ± 10 µm in mice treated with IMQ (Fig. 3; control). In mice treated with amthamine, JNJ-39758979 or the combination of both, the thickness of the epidermis was significantly lower than in the corresponding vehicle groups (Fig. 3). The thickness of the epidermis was lower in H4R–/– mice, but only by trend (Fig. 3).
Fig. 3. Amthamine and JNJ-39758979 attenuated imiquimod (IMQ)-induced epidermal thickening. IMQ strongly increased the epidermal thickness from approximately 20 ± 3 µm in mice treated with soft cream (control, far left) to 60 ± 10 µm in mice treated with IMQ. In mice treated with amthamine administered intraperitoneally (i.p.) and IMQ topically (histamine 2 receptor (H2R)+IMQ), the thickness of the epidermis was significantly lower than in mice treated with vehicle i.p. + IMQ topically (vehicle + IMQ). Also in mice treated with JNJ-39758979 i.p. + IMQ topically (histamine 4 receptor (H4R) + IMQ) or a combination of amthamine and JNJ-39758979 i.p. + IMQ topically (H2R+H4R+IMQ), the epidermis was significantly thinner than the epidermis of the corresponding vehicle groups (vehicle + IMQ). The epidermal thickness of wild-type (referred as “vehicle + IMQ”) and H4R–/– mice (referred as “treatment + IMQ”) was not significantly different. *p < 0.05 Mann–Whitney test. n = 6 BALB/c mice treated with amthamine and n = 7 BALB/c mice all other treatment groups; n = 6 wild-type and n = 6 H4R–/– mice. All drugs were given in a dosage of 20 mg/kg i.p. twice daily and were compared with treatment with vehicle (aqua ad injectionem, twice daily i.p.).
Amthamine and JNJ-39758979 reduced dermal lymphocytic cell infiltrates
In the skin of mice treated topically with soft cream, no lymphocytic infiltration was visible, whereas mice treated with IMQ showed a marked dermal infiltration of lymphocytic cells (Fig. 4A, B). In the skin of mice treated with the H2R agonist amthamine plus IMQ, lymphocytes persisted in the dermal layer, but the infiltration appeared to be less severe than in mice treated with vehicle plus IMQ (Fig. 4C, E). In the skin of mice treated with the combination of amthamine and JNJ-39758979 plus IMQ, lymphocytic infiltration of the dermis could only rarely be detected (Fig. 4H), whereas in the corresponding vehicle group a marked lymphocytic infiltration was again visible (Fig. 4G). In H4R–/– mice, lymphocytes persisted in the dermal layer, but infiltration appeared to be less severe than in wild-type mice (Fig. 4I, J).
Fig. 4. Imiquimod (IMQ)-induced dermal lymphocytic cell infiltrates. Representative histological pictures of lymphocytic cell infiltrates (asterisks) in the dermis. The treatment of mice using a combination of JNJ-39758979 and amthamine reduced lymphocytic cell infiltrates (asterisks) in the dermis. Skin of mice was treated topically with (A) soft cream or (B–J) IMQ for 10 consecutive days. (A) Skin of mice treated topically with soft cream. No lymphocytic infiltration (score (–)) was visible. (B) Skin of mice treated with IMQ topically. Lymphocytic infiltration (++++) is visible. (C, E, G) Skin of mice treated with vehicle administered intraperitoneally (i.p.) + IMQ topically. Lymphocytes (++++) persisted in the dermal layer (asterisks). (D) Skin of a mouse treated with amthamine i.p. + IMQ topically. Lymphocytes (+++) persisted in the dermal layer (asterisks), but infiltration appeared to be less severe than in mice treated with vehicle i.p. + IMQ topically (C). (F) Skin of a mouse treated with JNJ-39758979 i.p. + IMQ topically. Lymphocytes (+++) persisted in the dermal layer (asterisks), but infiltration appeared to be less severe than in mice treated with vehicle i.p. + IMQ topically (E). (H) Skin of a mouse treated with amthamine + JNJ-39758979 i.p. + IMQ topically. Lymphocytic infiltration (+) of the dermis could only rarely be detected. All drugs were given in a dosage of 20 mg/kg i.p. twice daily and were compared with treatment with vehicle (aqua ad injectionem, twice daily i.p.). (I) Skin of a wild-type mouse treated with IMQ topically. Lymphocytes (++++) persisted in the dermal layer (asterisks), infiltration appeared to be more severe than in H4R–/– mice (+++, J). Bars represent 50 µm.
H4R blockade reduces the number of lymph node cells
IMQ significantly increased the number of skin-draining lymph node cells compared with mice treated with soft cream (Fig. 5). The IMQ-induced increase was significantly lower in mice treated with H4R antagonist, but not in mice treated with H2R agonist compared with the corresponding vehicle groups (Fig. 5). Combination of JNJ-39758979 with H2R agonist clearly reduced the number of skin-draining lymph node cells compared with mice treated with vehicle (Fig. 5). In H4R–/– mice, the total number of skin-draining lymph node cells was significantly lower than in wild-type mice.
Fig. 5. Histamine 2 receptor (H2R) stimulation and histamine 4 receptor (H4R) blockade diminished imiquimod (IMQ)-induced increase in the cell number of skin-draining lymph nodes. IMQ greatly increased the total number of cells in the axillary lymph nodes. This increase was significantly reduced by amthamine (H2R), JNJ-39758979 (H4R) or the combination of both (H2R + H4R) compared with the corresponding vehicle (vehicle + IMQ) groups. Also, in H4R–/– mice, a significantly lower number of draining lymph node cells were detected compared with wild-type mice (named “vehicle” in the graph). *p < 0.05 Mann–Whitney test. n= 6 BALB/c mice treated with amthamine and n= 7 BALB/c mice all other treatment groups; n= 6 wild-type and n= 6 H4R–/– mice. All drugs were given in a dosage of 20 mg/kg administered intraperitoneally (i.p.) twice daily and were compared with treatment with vehicle (aqua ad injectionem, twice daily i.p.).
H4R blockade and H2R stimulation alone reduced pro-inflammatory cytokines in lesional skin
Significantly increased levels of Th17 cytokines IL-17A and IL-23 and of the Th1 cytokines (TNF-α and IL-33) were detected in the IMQ-treated skin at protein level compared with mice treated with soft cream (data not shown). The level of TNF-α was not significantly influenced by any treatment, and not reduced in H4R–/– mice compared with the corresponding vehicle groups or wild-type mice, respectively (Fig. S2A). The increase in IL-33 and IL-23 was diminished in amthamine and JNJ-39758979-treated mice, but not in mice treated with the combination of both drugs (Fig. S2B, D). The content of IL-17A in the skin was only significantly lowered by H2R agonist (Fig. S2C). In H4R–/– mice, a significantly lower amount of IL-17A and IL-23 was measured compared with wild-type mice, whereas IL-33 did not differ between groups (Fig. S2B–D).
Topical application of IMQ cream induces an inflammatory response in the skin that mimics psoriasis, with similar immunological pathways and cell types involved (9, 10). Since its introduction in 2009 by van der Fits et al. (10), it has become a widely used standard mouse model to mimic psoriasis and its associated itch.
Clinical cutaneous manifestations of psoriasis are diverse, but a hallmark of psoriasis is epidermal thickening and scaling (10). The H4R antagonist JNJ-39758979, as well as the H2R agonist amthamine, clearly reduced the scaling response and thickening. In a previous study, we showed that histamine induces proliferation of keratinocytes via activation of H4R (4). H2R ligands, in contrast, did not influence the proliferation of keratinocytes (4). In this study, however, the H2R agonist reduced IMQ-induced scaling and epidermal thickening. In line with this, topical treatment with an H2R antagonist stimulated epidermal proliferation after barrier disruption in mice (12). Thus, H2R most likely does not affect keratinocytes directly, but might influence proliferation-modulatory extracellular signals, such as the cytokine network in the local microenvironment.
Sakai et al. (13) reported that IMQ induces increased scratching behaviour in mice compared with vehicle cream, indicating that the IMQ-induced psoriasis model seems to be useful for the investigation of psoriatic itch. Interestingly, the combination of amthamine and JNJ-39758979 completely abolished spontaneous scratching behaviour in the IMQ model in the current study. Several studies in rodents and humans emphasize the role of H4R in histamine- and allergen-induced itch (14–18). Involvement of H2R in itch transmission, in contrast, is generally believed to be only marginal if at all (19). H2R agonists do not induce itch in humans or mice, and H2R antagonists do not inhibit histamine- or allergen-induced itch (15, 20, 21). However, in this study, the combination of an H2R agonist and an H4R antagonist almost completely abolished scratching behaviour, whereas the H4R antagonist alone did not. Whether itch response was because of the reduced skin score, or vice versa, the cause of reduced clinical score is not clear. In patients with psoriasis, there is not necessarily a correlation between the severity of the disease and the intensity of pruritus (22). The role of histamine in the pathogenesis of itch in psoriasis is unclear. Two studies showed that histamine levels in psoriatic skin were higher than those in healthy skin (2, 3). A more recent study, in contrast, found similar histamine levels in skin from healthy subjects and in psoriatic skin (23). Sakai et al. (13) reported that the expression level of histamine in IMQ-treated murine skin peaked on day 2 of IMQ treatment and returned to the basal level on day 3–7. Interestingly, JNJ-39758979 significantly reduced the scratching behaviour only on days 3 and 6 of IMQ treatment, which might be related to the histamine level in the skin. Both, H2R and H4R are expressed on murine dorsal root ganglia (DRG) and, for H4R at least, it was shown that stimulation of the receptor leads to Ca2+ influx in DRG (14). Thus, the anti-pruritic action of H4R antagonist might be a direct effect on sensory neurones.
Patients with psoriasis, as well as IMQ-treated mice, show increased production of the pro-inflammatory cytokines TNF-α and IL-33, as well as Th17 cytokines IL-17A and IL-23 (9-11, 24, 25), as confirmed in the current study. IL-23 induces IMQ- and psoriasis-associated epidermal hyperplasia and indirectly stimulates keratinocyte proliferation (24, 25). Increased levels of IL-17 mRNA and T cells producing IL-17 are found in psoriatic tissues (26). Mommert et al. (27) showed that activation of H4R stimulates Th17 cells to release enhanced amounts of IL-17, indicating that H4R leads to an exacerbation of inflammatory skin diseases, such as psoriasis. Inhibition of H4R and stimulation of H2R, however, decreased the IL-33 level in the skin. IL-33 levels are increased in skin lesions of patients with psoriasis, and it has been shown that psoriatic keratinocytes secrete this pro-inflammatory cytokine (28). The interaction of IL-33 with neutrophils and mast cells leads to a skin inflammation in mice that is similar to psoriatic plaques in humans (29). However, skin inflammation in the IMQ model did not differ between wild-type and IL-33–/– mice (30). In line with this, in the current study skin lesions were ameliorated in mice treated with a combination of amthamine and JNJ-39758979, even though the content of IL-33 was not decreased in the skin. Thus, reduced levels of the cytokines tested is probably not responsible for the anti-inflammatory effect seen by the treatment with amthamine plus JNJ-39758979.
Cell numbers of skin-draining lymph nodes were significantly reduced by amthamine and JNJ-39758979, underlining the reduction in the inflammatory response with these drugs. It was shown that topical application of IMQ yields systemic effects due to unintended oral uptake (31). JNJ-39758979 and the combination of amthamine and JNJ-39758979 could lower this reaction.
At sites of inflammation, Th1 and Th2 cells play important roles. It has been shown that both Th1- and Th2-type responses are negatively regulated by H2R through the activation of different biochemical intracellular signals. Mice lacking H2R showed an upregulation of both Th1 and Th2 cytokines, indicating an anti-inflammatory role of H2R (32). In human blood cells stimulation via H2R leads to an up-regulation of IL-10 and downregulation of IL-12. Antigen-presenting cells that are activated via H2R show a suppression of antigen presentation and lead to the formation of IL-10-producing T cells (reviewed in (33)). Also, pro-inflammatory responses to Toll like receptor (TLR) agonists are suppressed by histamine through H2R (34). This is interesting, as IMQ acts (at least partly) via activation of TLR7/8. Moreover, patients with chronic inflammation of the gut (Crohn’s disease) display dysregulated expression of histamine receptors, with diminished anti-inflammatory effects associated with H2R signalling (35). In summary, activation of H2R displays mainly anti-inflammatory properties that might also be beneficial in the context of psoriasis. Systemically, H2R agonist might not be well tolerated in humans, due to gastrointestinal or cardiac side-effects; however, topical application might be an option for psoriatic patients. H2R agonists have not been tested in psoriatic patients to date, but H2R antagonists have. They are not effective in the treatment of psoriasis, and some studies even report a worsening of the disease within the first month after treatment with H2R antagonists (36–39). Kristensen et al. (36) reported that the worsening seen after the first month of treatment with ranitidine was principally manifested as itching and increased redness. Interestingly, in the current study, amthamine significantly reduced the itch response and erythema within the first days of treatment, but not at a later time-point.
An H4R antagonist phase 2 clinical trial has been completed in patients with moderate-to-severe plaque-type psoriasis (40). The H4R antagonist was able to reduce the PASI score by a greater extent than placebo, but it did not meet the predefined success criterion (40). Thus, as in the mouse model, and in humans, blockade of H4R at least partially, inhibited the inflammatory response.
The results of the current study are contrary to those of Kim et al. (8), who showed that the H4R agonist 4-methylhistamine alleviated psoriatic symptoms in a dose-dependent fashion in the IMQ model. However, despite the relatively low affinity of 4-methylhistamine at H2R, it shows considerable in vivo H2R agonistic activities at dosages > 3 mg/kg i.v. in rodents (41). Reduction in skin lesions and epidermal thickening were seen at dosages of at least 20 mg/kg/day for 10 consecutive days, in a study by Kim et al. (8). Thus, the anti-inflammatory properties of 4-methylhistamine might be due to agonistic activity at H2R rather than agonistic activity at H4R.
In conclusion, this study found that combined treatment with an H2R agonist and H4R antagonist ameliorated skin inflammation and pruritus in a mouse model of psoriasis.
The authors thank the Deutsche Forschungsgemeinschaft and Janssen Research & Development, LLC. for supporting this study. We further thank Saskia Rübke for the preliminary work she performed for this study.
This study was supported by the Deutsche Forschungsgemeinschaft (DFG RO4649/1-1 and DFG RO4649/1-2). Funding for this research, in addition to JNJ-39758979, was provided by Janssen Research & Development, LLC. We acknowledge support by the German Research Foundation and the Open Access Publication Fund of the Freie Universität Berlin.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize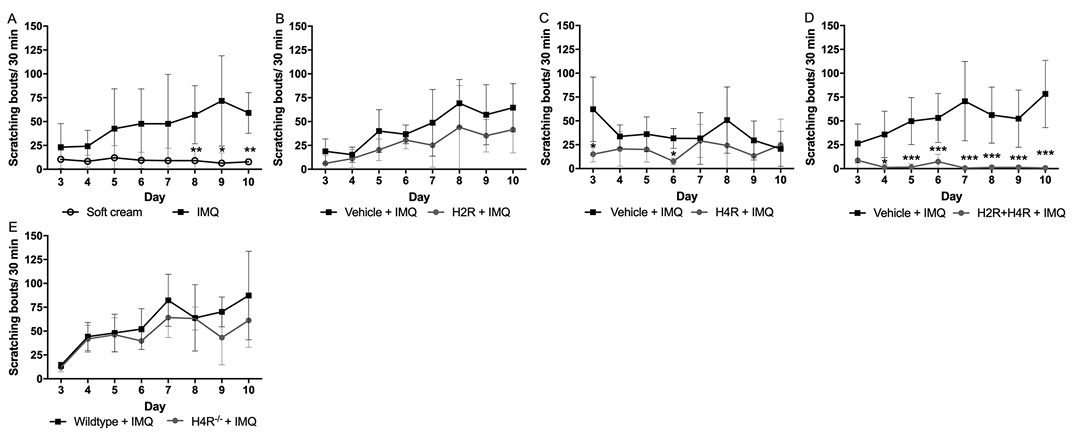 Click to show fullsize
Click to show fullsize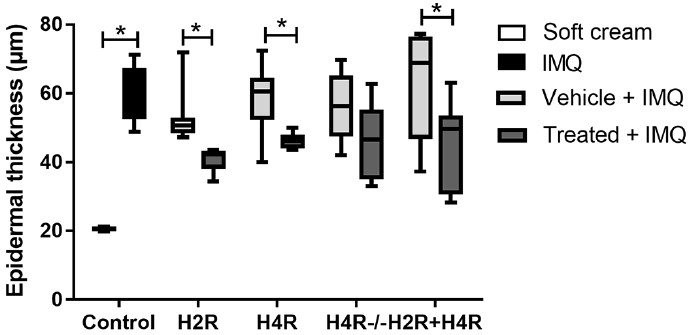 Click to show fullsize
Click to show fullsize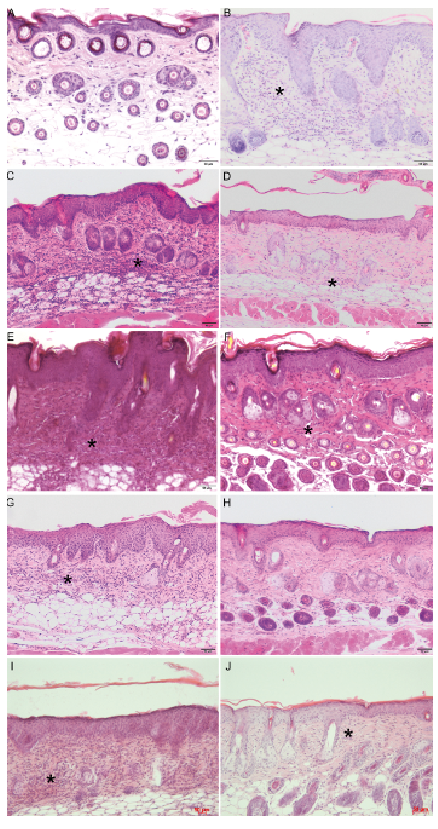 Click to show fullsize
Click to show fullsize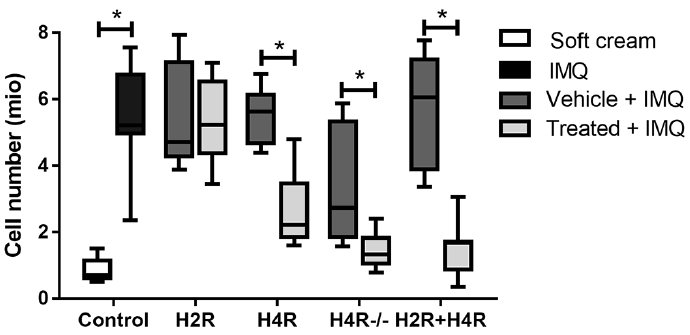 Click to show fullsize
Click to show fullsize