


1Department of Dermatology, 3Department of Transfusion Medicine and 4Division of Rheumatology, Department of Internal Medicine III, Medical University of Vienna, and 2Core Facility Cell Imaging and Ultrastructure Research, University of Vienna, Vienna, Austria
Cathelicidins have been reported to inhibit human papillomavirus infection in vitro; however, nothing is known about their activity in vivo. In this study, experimental skin infection with Mus musculus papillomavirus 1 resulted in robust development of cutaneous papillomas in cyclosporine A-treated C57BL/6J mice deficient for the murine cathelicidin-related antimicrobial peptide (CRAMP), in contrast to wild-type controls. Analysis of the underlying mechanisms revealed moderate disruption of virion integrity and lack of interference with viral entry and intracellular trafficking by a synthetic CRAMP peptide. Differences in the immune response to Mus musculus papillomavirus 1 infection were observed between CRAMP-deficient and wild- type mice. These included a stronger reduction in CD4+ and CD8+ T-cell numbers in infected skin, and lack of Mus musculus papillomavirus 1-specific neutralizing antibodies in response to cyclosporine A in the absence of endogenous CRAMP. CRAMP has modest direct anti-papillomaviral effects in vitro, but exerts protective functions against Mus musculus papillomavirus 1 skin infection and disease development in vivo, primarily by modulation of cellular and humoral immunity.
Key words: antimicrobial peptide; cathelicidin; CRAMP; MmuPV1; skin papilloma; antiviral mechanisms.
Accepted Dec 17, 2020; Epub ahead of print Dec 22, 2020
Acta Derm Venereol 2021; 101: adv00367.
doi: 10.2340/00015555-3733
Corr: Alessandra Handisurya, Department of Dermatology, Medical University of Vienna, Waehringer Guertel 18–20, AT-1090 Vienna, Austria. E-mail: alessandra.handisurya@meduniwien.ac.at
Key words: antimicrobial peptide; cathelicidin; CRAMP; MmuPV1; skin papilloma; antiviral mechanisms.
Cathelicidins are host defence (antimicrobial) peptides. This study demonstrates that mice, which lack the murine cathelicidin-related antimicrobial peptide in their skin and immune cells, consistently developed large papillomas on the skin after infection with a mouse papillomavirus and concomitant immunosuppression. In contrast, the presence of cathelicidin-related antimicrobial peptide in control mice was protective against development of skin papillomas. The cathelicidin-related antimicrobial peptide-mediated protection against papillomavirus skin infection and associated disease was mediated by modulation of the host’s immune system. These results indicate cathelicidins as important targets for future anti-papillomaviral compounds.
Cathelicidins, such as human LL-37 and the murine orthologue cathelicidin-related antimicrobial peptide (CRAMP), are endogenous host defence peptides with broad antimicrobial and immunomodulatory functions. They are produced by many different cells, including neutrophils, leukocytes, and epithelial cells, and are expressed in numerous tissues, such as skin, intestine, and bone marrow (1, 2). In healthy human skin, expression of LL-37 is constitutively low, but can be induced by inflammatory or infectious stimuli (3). LL-37 has also been detected in human papillomavirus-induced genital and skin warts (4). Besides their role in the innate and adaptive immune defence against bacterial and fungal infections, cathelicidins demonstrate direct effects in vitro against several viruses, such as respiratory syncytial- (RSV), influenza A- (IAV), varicella zoster- (VZV), herpes simplex- (HSV), and vaccinia viruses (VV), through disruption of virions, interference with virus-cell binding, and inhibition of viral replication (5–9). Furthermore, exogenous administration of cathelicidins protected against virus infection in vivo and/or reduced disease activity and viral replication, as shown for RSV and IAV (6, 10). In mice deficient for endogenous CRAMP (Cnlp-null or CRAMP-knock-out (KO) mice), which lacked expression in neutrophil granules and epidermal keratinocytes (11), RSV-, VV- or HSV-induced disease severities were more pronounced and viral loads higher compared with wild-type (WT) mice, which endogenously expressed CRAMP (7, 8, 10).
Papillomaviruses (PVs) are non-enveloped, double-stranded DNA viruses that target epithelia of humans and many vertebrate species for infection with high species-specificity. PV infections are mostly asymptomatic and transient, but can lead to development of benign papillomas (warts) of skin and mucosa in their respective host. Persistent infections with certain oncogenic high-risk mucosal human PVs are established causes of anogenital and oropharyngeal cancers in humans. To date, antiviral activity of cathelicidins against infection of epithelial cells with PV-derived pseudovirions (PsVs), which display properties similar to native, infectious virions, has been demonstrated in vitro (12). While this suggests a possible protective effect of cathelicidins against PV infections and associated diseases, in vivo investigations have not yet been performed.
The murine PV, Mus musculus papillomavirus 1 (MmuPV1), infects laboratory mice and induces outgrowth of skin papillomas in severely immunocompromised mice. In contrast, infection of immunocompetent mice is generally asymptomatic and transient, as PV infection and associated disease are controlled and cleared by the cellular immunity (13, 14).
The aim of the current study was to investigate the effects of the murine cathelicidin CRAMP against MmuPV1 skin infection and associated skin papillomatosis in genetically modified C57BL/6J mice deficient for CRAMP. In addition, potential antiviral mechanisms of CRAMP, such as disruption of particle integrity, inhibition of virus entry into and intracellular trafficking in infected keratinocytes, or modification of virus infection through modulation of immunity, were addressed.
Ethics statement
The animal experiments were approved by the ethics committee of the Medical University of Vienna, Austria, and the Austrian Federal Ministry of Science and Research (BMWFW-66.009/0209-WF/V/3B/2016, BMWFW-66.009/0208-WF/V/3B/2016) and performed in full compliance with the institutional guidelines.
Mus musculus papillomavirus 1 skin infection model
Infectious MmuPV1 virions were purified from papillomatous tissue extracts derived from athymic nude mice, as described previously (15). Female C57BL/6J mice, aged 5–6 weeks, deficient for CRAMP in skin (Fig. S1), bone marrow, and neutrophil granules (11), (n = 16) and WT controls (n = 16) were obtained from Charles River Laboratories Inc. (Wilmington, MA, USA). Experimental infection was performed with native MmuPV1 virions at 1×1012 per site on the skin of the tails (16). Systemic immunosuppression was performed by subcutaneous administration of cyclosporine A (CsA) (Sandimmune® Inject., Novartis, Basel, Switzerland) at a dose of 75 mg/kg body weight starting one week prior to infection 5 times per week for a total of 6 weeks (13). Mice were followed for the development of skin papillomas and at indicated time-points sera were obtained and skin tissues procured from the inoculation sites.
Determination of MmuPV1-E1^E4 spliced transcripts, copy numbers and L1 capsid protein
Crude skin tissue extracts were prepared, as described previously (15). Total RNA and genomic DNA were purified from the same skin tissue sample using TRI reagent (Sigma-Aldrich, St Louis, MO, USA) according to the manufacturer’s instructions. MmuPV1-E1^E4 spliced transcripts, a marker for infectivity and viral transcription, and viral genome copy numbers were determined by real-time PCR, employing previously published primers and probes (15). MmuPV1-E1^E4 levels were normalized to the endogenous control ACTB (Thermo Fisher Scientific, Waltham, MA, USA) and calculated using the Comparative CT (ΔΔCT) method. The MmuPV1 copy numbers in the DNA samples were quantified by comparison with defined amounts of the MmuPV1 genome as standards after normalization to the housekeeping gene γ-actin (15,17). Detection of the major capsid protein L1 in the skin tissue extracts was performed by Western blot, employing a polyclonal rabbit immune serum raised against MmuPV1-L1 at a dilution of 1:1,000 (15).
Haematoxylin-eosin staining and immunohistochemistry
Skin tissue sections were stained with haematoxylin-eosin or used for immunohistochemistry employing following primary antibodies: rabbit polyclonal CRAMP (diluted 1:2,000) (Innovagen AB, Lund, Sweden), rat-anti-mouse CD4 (diluted 1:10) (BD Biosciences, San Jose, CA, USA) and rat-anti-mouse CD8 (dilution 1:100) (BioLegend, San Diego, CA, USA). The MmuPV1 L1/L2-specific polyclonal immune serum was generated (Eurogentec, Seraing, Belgium) by immunization of a New Zealand White rabbit with 0.1 mg MmuPV1 L1/L2 PsVs in a 4-dose regimen at week 0-2-4-8. Serum was drawn 4 weeks after the final boost, affinity-purified and employed for immunohistochemistry diluted 1:4,000. All images were digitalized using an Aperio slide scanner (Leica Biosystems, Nussloch, Germany) and immunopositive cells per mm2 skin tissue semi-quantitatively scored on anonymized samples using ImageJ.
Single-molecule RNA in situ hybridization
Detection of the MmuPV1 oncogene E6/E7 mRNA was performed, employing the RNAscope® MusPV-E6-E7 probe (Advanced Cell Diagnostics, Newark, CA, USA) according to the manufacturer’s instructions. RNA integrity was verified with the endogenous control probe Mm-PPIB and background staining evaluated, employing the negative control probe specific for the bacterial DapB gene. Brightfield images were acquired using the Aperio slide scanner (Leica Biosystems, Nussloch, Germany).
Peptide synthesis
The synthetic murine CRAMP peptide was commercially obtained (Bachem, Bubendorf, Switzerland) and the scrambled control peptide (LQPKVELQLKGIPGQEKKGNFRGLKGEKKQFIKI), containing the same amino acids in a randomly rearranged order, synthesized (JPT Peptide Technologies GmbH, Berlin, Germany).
Peptide-enzyme-linked immunosorbent assay
Peptide binding to native virions was assessed by peptide-enzyme-linked immunosorbent assay (ELISA) (18) and bound peptide-virion-complexes detected, employing the polyclonal rabbit immune serum recognizing MmuPV1-L1/L2.
Transmission electron microscopy analyses
To assess particle integrity 1×1011 MmuPV1 virions were co-incubated with either the CRAMP or the control peptide at final concentrations ranging from 10–100 µg/ml for 24 h, adsorbed onto glow-discharged copper grids, fixed and negatively stained with 1% uranyl acetate (19). Micrographs were taken at a magnification of 60,000× at 60 kV, employing a JEOL 1010 electron microscope (JEOL GmbH, Eching, Germany). A minimum of 3 grids were analysed for each condition by 3 independent individuals blinded to the experimental allocations.
S1 mouse keratinocytes were seeded on Aclar® discs (Electron Microscopy Sciences, Hatfield, PA, USA) at a density of 4 × 104 cells per disc, infected with 1×109 MmuPV1 virions and co-incubated with 50 µg/ml synthetic CRAMP or control peptide. Sixteen hours post-infection cells were fixed and embedded in Agar-100 resin (Agar Scientific Ltd, Stansted, UK). Ultrathin sections of embedded cell layers were mounted on copper grids and contrasted with uranyl acetate/lead citrate, prior to analyses at 120 kV employing a Libra 120 electron microscope (Carl Zeiss AG, Oberkochen, Germany). Acquisition of images was performed using a bottom stage digital camera, TRS (4 megapixel) and ImageSP professional software (Tröndle, Moorenweis, Germany).
Detection of Mus musculus papillomavirus 1-specific antibodies in mouse sera
MmuPV1-specific antibodies in the sera (dilution range 1:100 to 1:10,0000) of CRAMP-KO and WT mice were detected by particle-enzyme-linked immunoassay (particle-ELISA), employing native MmuPV1 virions as the antigen (20). MmuPV1-SEAP PsVs were obtained by co-transfection of 293TT cells with pMusheLL and a reporter plasmid encoding for secreted alkaline phosphatase (SEAP) (21). The presence of anti-MmuPV1 antibodies with neutralizing capacity in the mouse sera (dilution 1:100) was assessed, employing MmuPV1-SEAP PsVs (22). The IC50 titre is defined as the highest serum dilution showing at least 50% reduction in SEAP signal compared with the negative controls, mouse pre-immune sera or in the absence of immune sera.
Statistical analyses
Statistical analyses were performed using GraphPad Prism 7. Differences between groups were analysed using Mann–Whitney test or 1-way analysis of variance (ANOVA) at 95% confidence interval (95% CI). A p-value < 0.05 was considered statistically significant. Data represent mean ± standard deviation (SD).
Mus musculus papillomavirus 1 infection causes skin papillomas in cyclosporine A-treated cathelicidin-related antimicrobial peptide-deficient C57BL/6J mice
C57BL/6J mice deficient for endogenous CRAMP consistently (100% at each time-point; 10/10 at day 23 and 5/5 at day 35 post-infection) developed cutaneous papillomas on their tails after MmuPV1 infection and systemic CsA immunosuppression (Fig. 1a). The papillomas were large, with mean lengths 16.2 ± 7.4 mm on day 35 post-infection. Histological analyses of tissue specimens additionally verified the presence of papillomas (Fig. 1b). Only a subset (40% at each time-point; 4/10 at day 23 and 2/5 at day 35 post-infection) of CsA-treated, MmuPV1-infected WT C57BL/6J controls, in contrast, developed skin papillomas (Fig. 1c) and the lesions were much smaller at 6.5 ± 9.1 mm.
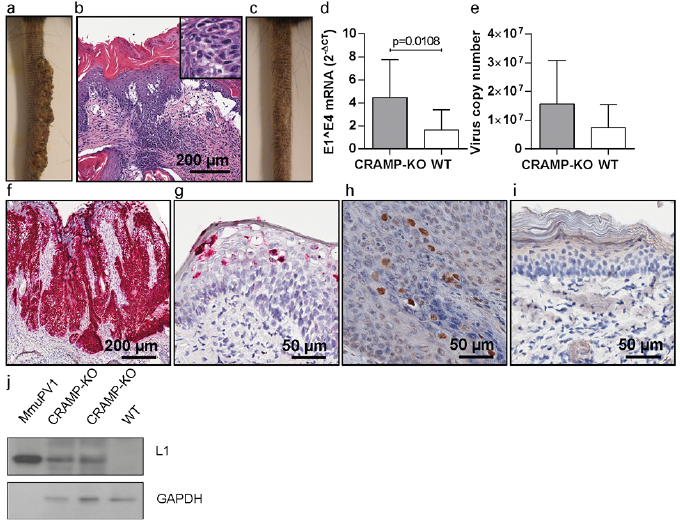
Fig. 1. Mus musculus papillomavirus 1 (MmuPV1)-induced skin papillomas in cyclosporine A (CsA)-treated cathelicidin-related antimicrobial peptide (CRAMP)-knock out (KO) and wild-type (WT) mice. (a) CsA-treated, MmuPV1-infected CRAMP-deficient C57BL/6J mouse on day 35 post-infection. (b) Haematoxylin-eosin (HE) staining of a skin papilloma, showing hyperkeratosis, acanthosis, elongation of the rete ridges. Koilocytes in the epithelium are depicted in the insert. (c) CsA-treated, MmuPV1-infected WT C57BL/6J mouse on day 35 post-infection. (d) Levels of MmuPV1-E1^E4 spliced transcripts and (e) MmuPV1 viral copy numbers in infected tail skin tissues of CRAMP-KO and WT mice. MmuPV1-E6/E7 oncogene mRNA in (f) the papillomatous lesions of MmuPV1-infected CRAMP-KO mice and (g) infected skin of MmuPV1-infected WT controls. MmuPV1-L1/L2 capsid proteins in the epidermal skin layers of (h) MmuPV1-infected CRAMP-deficient and (i) MmuPV1-infected WT mice. (j) MmuPV1-L1 major capsid protein in crude skin tissue extracts derived from infected CRAMP-KO and WT mice. Purified MmuPV1 virions served as positive control.
To assess viral gene expression, skin tissues were harvested from the infection sites, regardless of whether visible lesions had evolved. In the tissues of CsA-treated, MmuPV1-infected CRAMP-KO mice abundant MmuPV1-E1^E4 spliced transcripts were present and the levels were significantly higher than in the tissues derived from equally treated, infected WT controls (p = 0.0108) (Fig. 1d). Similarly, the mean viral copy numbers in the infected tissues were approximately twice as high in the CRAMP-KO compared with the WT mice (Fig. 1e) and detection of these viral markers correlated well with the macroscopic appearances of the papillomas (not shown). The viral contribution was further confirmed by abundantly expressed MmuPV1-E6/E7 mRNA throughout the papillomas of the CRAMP-KO mice, demonstrating increased transcriptional activity of the viral oncogenes in the absence of endogenous CRAMP (Fig. 1f). In contrast, E6/E7 mRNA in the keratinocytes was sparse in the WT mice (Fig. 1g). Expression of the MmuPV1-L1/L2 capsid proteins in keratinocytes was restricted exclusively to tissues of CRAMP-KO mice, but not WT controls (Fig. 1h–j). The results demonstrate the role of endogenous CRAMP in protection against MmuPV1 skin infection and manifestation of virus-induced disease in vivo.
The immunosuppressive effect of CsA was a prerequisite for papilloma development, as lesional outgrowth was not observed macroscopically and histologically during the 8-week follow-up period in non-CsA-treated, MmuPV1-infected CRAMP-deficient (0%; 0/6) and WT C57BL/6J mice (0%; 0/6) (Fig. S2). Non-CsA-treated CRAMP-KO and WT mice also did not express MmuPV1-E1^E4, E6/E7 mRNA and L1/L2 proteins in infected tissues, and only very low viral copy numbers, which could be attributable to residual, partially cleared inoculum, were detectable (data not shown). These findings correspond with the absence of skin papillomas, possibly due to rapid elimination of viral infection in the C57BL/6J strain, which is regarded as resistant to MmuPV1-induced skin papillomatosis even under severe immunosuppressed conditions (13, 14).
Synthetic cathelicidin-related antimicrobial peptide interacts directly with native Mus musculus papillomavirus 1 virions
To determine whether CRAMP can interact directly with MmuPV1, binding of a synthetic CRAMP peptide to native virions was assessed by ELISA. Binding to the CRAMP peptide was specific, dose-dependent and higher (p = 0.0004) compared with a scrambled control peptide (Fig. 2a), demonstrating that CRAMP could interact directly with native MmuPV1 particles.
Fig. 2. Cathelicidin-related antimicrobial peptide (CRAMP)-Mus musculus papillomavirus 1 (MmuPV1) virion interactions. (a) Binding of MmuPV1 virions to a synthetic CRAMP and a scrambled control peptide. One representative result out of 3 independent experiments is shown. (b) Electron micrographs showing damaged MmuPV1 virions, particle conglomerates and fragments in the presence of the synthetic CRAMP peptide and (c) intact particles in the presence of the scrambled control peptide (24-h exposure; final concentration 100 µg/ml). (d) Comparison of damaged virions after exposure to synthetic CRAMP or the control. (e) Thin-section electron micrographs showing native MmuPV1 virions attached to the keratinocyte surface, (f) bound in indentations of the cell membrane, (g) engulfed in intracellular vesicles in the presence of the synthetic CRAMP peptide (16-h time-point, final concentration 50 µg/ml). (h) Intravesicular virions in the keratinocyte in the presence of the control peptide (16-h time-point; final concentration 50 µg/ml).
Cathelicidin-related antimicrobial peptide affects the integrity of native Mus musculus papillomavirus 1 virions
To study the effect of CRAMP on particle integrity, native MmuPV1 virions were co-incubated with either the synthetic CRAMP or the scrambled control peptide, and the morphology of the viral particles was subsequently analysed by transmission electron microscopy. Exposure to the CRAMP peptide affected the structural integrity of the virions, as demonstrated by numerous broken particles with damaged membranes, particle conglomerates and fragments (Fig. 2b). Incubation with the control, in contrast, had no effect on the particle structure and the virions remained relatively intact (Fig. 2c), similar to control grids, where MmuPV1 virions were analysed in the absence of any peptide (not shown). Quantification analyses revealed that co-incubation of CRAMP resulted in a higher, albeit statistically not significant, percentage of damaged viral particles compared with the control at 8.4% vs 4.4%, respectively (p = 0.1000) (Fig. 2d). This effect was dose-dependent, as co-incubation with lower amounts of CRAMP resulted in lower numbers of defective particles (not shown).
Cathelicidin-related antimicrobial peptide does not impair virus entry into keratinocytes and intracellular trafficking
Next, the capability of CRAMP to interfere with virion-cell binding, virus entry and/or intracellular trafficking was assessed. Native MmuPV1 virions bound to the keratinocyte surface could readily be detected in the presence of the CRAMP peptide and were also found in slight indentations in the cell membranes, indicating virion uptake (Fig. 2e, f). Inside the keratinocytes, viruses were present in small vesicles up to 300 nm in length, that harboured one to several particles (Fig. 2g). This proved that binding, internalization and the initial steps of intracellular trafficking of MmuPV1 virions were not disturbed by CRAMP. Similarly, intracellular virus particles were also observed in the presence of equal amounts of control peptide (Fig. 2h).
Cyclosporine A differently affects CD4+ and CD8+ T-cells in Mus musculus papillomavirus 1-infected skin of cathelicidin-related antimicrobial peptide-knock-out and wild-type mice
Next, this study investigated the cellular infiltrate, particularly of CD4+ and CD8+ T-cells, which represent the major players responsible for PV control and elimination, in the infected skin tissues. In non-CsA-treated, MmuPV1-infected mice numerous CD4+ and CD8+ T-cells were generally present in the skin tissues, particularly of the CRAMP-deficient mice (Fig. 3a, b). Quantification analyses revealed that the mean CD4+ and CD8+ T-cell numbers were 6.9-fold and 1.5-fold higher, respectively, in non-CsA-treated, MmuPV1-infected CRAMP-KO mice compared with WT controls. In the presence of CsA, however, papilloma-bearing MmuPV1-infected CRAMP-KO mice had 2.5-fold lower mean CD4+ T-cell numbers per mm2 tissue compared with equally treated, MmuPV1-infected WT controls and 7.9-fold lower CD4+ numbers compared with non-CsA-treated, MmuPV1-infected littermates (Fig. 3a). CsA treatment, in contrast, did not reduce CD4+ counts in MmuPV1-infected WT mice, but resulted in 2.2-fold higher numbers compared with non-CsA-treated littermates. The mean CD8+ T-cell numbers were equally low in the skin tissues of CsA-treated, MmuPV1-infected CRAMP-KO and WT mice (Fig. 3b). CsA administration significantly reduced the mean CD8+ T-cell numbers by 30.6-fold in MmuPV1-infected CRAMP-KO mice compared with non-CsA-treated, MmuPV1-infected littermates. Similarly, CsA-treated, MmuPV1-infected WT mice had 37.4-fold lower mean CD8+ T-cell numbers compared with non-CsA-treated, infected littermates. This indicated that CsA-immunosuppression affected CD4+ rather than CD8+ T-cells in MmuPV1-infected mice deficient for endogenous CRAMP.
Fig. 3. CD4+ and CD8+ T-cell infiltrates in skin tissues and Mus musculus papillomavirus 1 (MmuPV1)-specific neutralizing antibodies in sera of MmuPV1-infected cathelicidin-related antimicrobial peptide (CRAMP)-knock out (KO) and wild-type (WT) mice. (a) Quantification of mean CD4+ T-cell numbers and (b) CD8+ T-cell numbers per mm2 skin tissue of CsA-treated, MmuPV1-infected CRAMP-KO and WT and non-CsA-treated, MmuPV1-infected CRAMP-KO and WT mice. (c) Mean IC50 titres of MmuPV1-specific neutralizing antibodies (as determined by pseudovirion neutralization assay) in sera derived from CsA-treated, MmuPV1-infected CRAMP-KO and WT and non-CsA-treated, MmuPV1-infected CRAMP-KO and WT mice.
Cyclosporine A-treated cathelicidin-related antimicrobial peptide-knock-out mice fail to mount Mus musculus papillomavirus 1-specific antibodies with neutralizing capacity
Next, this study investigated the effects on the humoral immune response induced by endogenous CRAMP after MmuPV1 infection. Non-CsA-treated as well as CsA-treated CRAMP-KO and WT mice were able to mount a MmuPV1-specific antibody response after infection, as shown by virus particle-ELISA (Fig. S3). The functionality of the MmuPV1-specific antibodies, however, differed significantly between the individual experimental groups. Antibodies specific for neutralizing epitopes in the papillomaviral capsid proteins, which represent the antibody group capable of mediating protection from viral infection (23), were readily detectable in non-CsA-treated, MmuPV1-infected mice (Fig. 3c) with significantly higher mean IC50 titres in the CRAMP-KO compared with the WT mice (p = 0.0005). Intriguingly, in the vast majority (80%; 8/10) of the CsA-treated, MmuPV1-infected CRAMP-KO mice, neutralizing antibodies were undetectable, and in the remaining 2 mice (20%) levels were low. In contrast, neutralizing antibodies were present in sera of CsA-treated, MmuPV1-infected WT mice at higher mean IC50 titres that were relatively comparable to the titres observed in non-CsA-treated, MmuPV1-infected littermates. Uninfected mice lacked seroresponses (not shown).
This study demonstrates that endogenous CRAMP is protective against MmuPV1-induced skin papillomatosis in vivo, as CRAMP-deficient C57BL/6J mice consistently developed large papillomas after systemic treatment with CsA. Abundantly expressed viral markers in the papillomas were indicative for active transcription and virion production. The C57BL/6J strain was previously reported to be resistant to MmuPV1-induced papillomatosis due to immunological factors specific to this strain, in contrast to mice on a different genetic background (13, 14). To overcome the strain-specific resistance to virus-induced skin papillomatosis, systemic CsA was administered to lower T-cell activity and higher amounts of MmuPV1 virions (1×1012 as opposed to 4.2×1010 per site in the previous study (13)) were used for infection. The latter would explain why some of the CsA-treated C57BL/6J WT mice developed lesions, albeit small in size.
Previous reports have shown direct mechanisms how cathelicidins, mostly human LL-37, exert antiviral activity against a variety of viruses (5–9, 24, 25). PVs lack a viral envelope, hence, direct killing through permeabilization of the envelope layer is ruled out as the operative mechanism. While very modest effects of a synthetic CRAMP peptide on virion integrity were demonstrated, this mechanism alone seems insufficient to explain the observed in vivo protection of WT mice against MmuPV1-induced skin papillomatosis. Internalization of virions into murine keratinocytes and intracellular trafficking, including deposition into vesicles, were not inhibited by the CRAMP peptide, although we have not investigated the fate of the viral particles, as an impact of the released peptide on the transition of the virus from the endosome to the nucleus seems unlikely.
In general, cytotoxic CD8+ T-cell responses are responsible for the control and clearance of many viral infections and for the elimination of virus-infected cells, whereas certain CD4+ T-cells modulate cytotoxic T-cell activation via production of antiviral cytokines and release of cytotoxic molecules. In addition, protective antiviral cytotoxic CD4+ T-cell responses have been observed during a multitude of virus infections, including herpes-, cytomegalo-, and lymphochoriomeningitis viruses (26, 27), as well as in MmuPV1-infected C57BL/6J mice (13). The current study demonstrated strong CD4+ and CD8+ T-lymphocytic infiltration in the skin of CRAMP-expressing WT and CRAMP-deficient KO mice after MmuPV1 infection, which presumably prevented virally-induced papilloma outgrowth. In the presence of systemic CsA, which inhibits T-lymphocyte activation by targeting calcineurin and the mitogen-activated protein kinases JNK and p38 within the interleukin 2 pathway (28), differential T- and B-cell responses were observed in the absence or presence of CRAMP.
In CsA-treated CRAMP-expressing WT mice CD8+ T-cells were reduced, whereas CD4+ T-cells were up-regulated. Given the possible cytotoxic capacity of the CD4+ T-cells, this T-cell subset may suffice to control viral infection and prevent lesional outgrowth, even in the absence of CD8+ T-cells. In this line, the CsA-treated WT mice which had fewer CD4+ T-cells were the ones that developed small papillomas. In the infected skin tissues of CRAMP-deficient mice, CsA significantly reduced both CD4+ and CD8+ T-cells. It is conceivable that, in the absence of endogenous CRAMP, the remaining CD4+ T-cells might not be sufficient in numbers to protect against papillomatosis, for instance in exerting the required cytotoxic effector functions for viral elimination or in providing sufficient help to the CD8+ T-cells. Alternatively, T-cell function might be altered in the CRAMP-KO mice in response to CsA, thus impairing efficient suppression of viral infection and papillomatosis.
Next to the cytotoxic activity of T-cells, neutralizing antibodies are important in viral defence, in particular during the state prior to viral entry into the target cell. The current study showed striking differences in the B-cell responses to MmuPV1 infection between WT and CRAMP-KO mice with the effect of CsA immunomodulation. In general, PV-specific antibodies induced either after vaccination or after infection are not active against established disease, but prevent new infection. While we do not believe that the neutralizing antibodies generated in this study in the mice after MmuPV1 infection play a decisive role in protecting against virus infection and disease, due to the latency after which antibodies are generated, the complete absence of neutralizing antibodies in the CsA-treated CRAMP-KO mice was striking, and suggested that the absence of endogenous CRAMP might impair T-cell mediated B-cell maturation and function in response to CsA. Another possible explanation is that antibody-dependent cytotoxicity is impaired in the CsA-treated CRAMP-KO mice. In this scenario, the lack of (functionally active) antibodies would prevent recognition and subsequent elimination of virus-infected cells by cytotoxic T-cells. The presence of MmuPV1-specific antibodies in both CsA-treated KO and WT mice argues against antibodies as the effective key players in preventing MmuPV1-induced papillomatosis, but rather points to the presence of cytotoxic CD4+ T-cells and, indirectly, to a role for CRAMP in T-cell regulation and/or activation. We cannot exclude that other cellular players might also be involved in in vivo protection of CRAMP against papillomaviral infection, such as skin-infiltrating plasmacytoid dendritic cells and/or Langerhans cells. In the absence of CRAMP, antigen-presentation may be impaired in the CRAMP-KO mice, especially during a subdued immunological state; for example, by CsA.
In conclusion, this study showed that protection against MmuPV1-induced skin papillomatosis in vivo is mediated by CRAMP. Absence of CRAMP, together with CsA immunosuppression, affects both cytotoxic T-cell activity and specific antibody production, indicating cathelicidins as important targets for future anti-papillomaviral compounds.
This research was supported by the Austrian Science Fund (FWF P29619-B30) and the Medical-Scientific Fund of the Mayor of Vienna (BGM 17019). Electron microscopy sample preparation and analyses of keratinocytes were performed at the Core Facility Cell Imaging and Ultrastructure Research, University of Vienna – member of the Vienna Life-Science Instruments (VLSI). The authors thank Roland Tresky, Ruth Dingelmaier-Hovorka, and Ulrike Mann for their excellent technical assistance, Wolfgang P. Weninger, MD, for helpful discussions and John T. Schiller, PhD, for valuable suggestions and critical reading of the manuscript. S1 mouse keratinocytes and 293TT cells were kindly provided by John T. Schiller, PhD, and Patricia M. Day, PhD, National Institutes of Health, National Cancer Institute, Bethesda, MD, USA.
The authors have no conflicts of interest to declare.


 Click to show fullsize
Click to show fullsize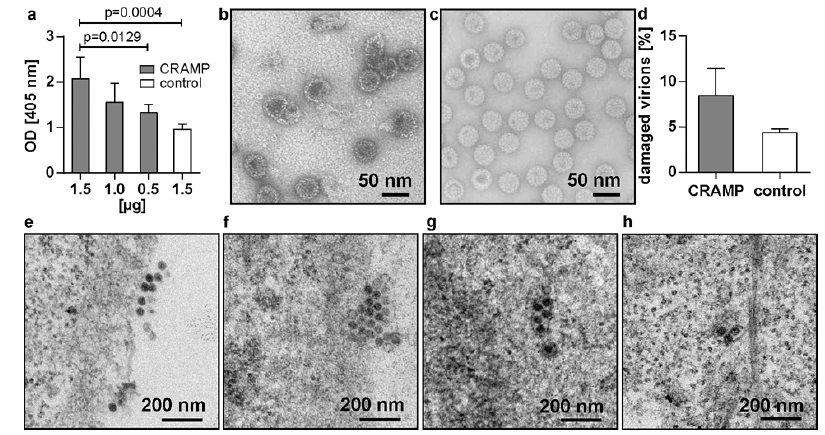 Click to show fullsize
Click to show fullsize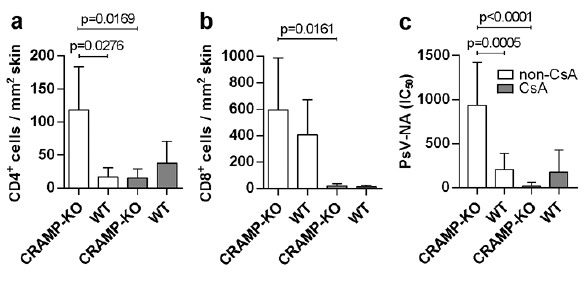 Click to show fullsize
Click to show fullsize