


Thilo Gambichler1, Lena Reininghaus1, Marina Skrygan1, Hans-Joachim Schulze2, Jörg Schaller3, Chiara Colato4 and Giampiero Girolomoni5
1Department of Dermatology, Ruhr-University Bochum, Gudrunstrasse 56, DE-44791 Bochum, 2Fachklinik Hornheide, Skin Cancer Centre, Department of Dermatology and Dermato-Histo-Pathology, Münster, 3Dermatopathology Duisburg, Duisburg, Germany, 4Section of Pathology, Department of Pathology and Diagnostics, and 5Section of Dermatology and Venereology, Department of Medicine, University of Verona, Verona, Italy. E-mail: t.gambichler@klinikum-bochum.de
Accepted Jan 14, 2016; Epub ahead of print Jan 18, 2016
Mid-dermal elastolysis (MDE) and anetoderma (AN) are rare, closely-related, skin disorders of the elastic tissue, which are histopathologically characterized by loss of elastic fibres in the dermis (1, 2). In MDE, most commonly observed skin changes include patches of well-circumscribed fine wrinkles (type I) and areas of perifollicular papular protrusions (type II). Rarely, MDE may also occur with persistent reticular erythema and wrinkling (type III). In AN, localized macular lesions consisting of flaccid or herniated saclike skin are observed with or without inflammatory signs (1–3). Both MDE and AN are benign conditions. However, they can cause significant cosmetic impairment and disfigurement. The pathogenesis of MDE and AN may reflect an imbalance of dermal elastin turnover, as evidenced by elevated levels of several members of the matrix metalloproteinases (MMPs) without an equivalent increase in tissue inhibitors of metalloproteinases (2–7). A possible role of macrophages has been described, particularly in AN (1, 8). We have previously shown, in a patient with MDE, that not only increased elastolytic activity (e.g. MMP9 up-regulation), but also elastin renewal due to altered lysyl oxidase-like 2 (LOXL2) enzyme expression may contribute to the pathogenesis of MDE (7). More research is needed, especially with regard to the LOX/LOXL family, fibrillins, desmosines and fibulins. The latter protein family is composed of long and short fibulins, which bind the monomeric form of elastin (tropoelastin) and are involved in various aspects of elastic fibre development (9). We report on the protein expression of fibulin-4 and fibulin-5 in patients with MDE and AN.
MATERIALS AND METHODS
We investigated archived lesional formalin-fixed and paraffin-embedded tissue of 14 patients with MDE (mean age ± standard deviation (SD) 41.4 ± 12.8 years; 10 women (71.4%) and 4 men (28.6%)) and 9 patients with idiopathic AN (mean age ± SD 39.6 ± 18.2 years; 8 women (88.9%) 1 man (11.1%)). Diagnoses were based on history, clinical picture and histopathology, including elastic van Gieson stains. Moreover, skin tissue of 10 healthy sex-/age-matched subjects was investigated as controls (mean age ± SD 42.5 ± 13.3 years; 8 women (80%) and 2 men (20%)). The study was approved by the ethics review board of the Ruhr-University Bochum (#4222-12).
Immunohistology for fibulin-4 and fibulin-5 was performed as described previously (5, 6). The following antibodies were used: rabbit monoclonal fibulin-4 (EPR684(2)) antibody at 1:250 dilution and pH 6 (Abcam plc, Cambridge, UK); mouse monoclonal fibulin-5 (1G6A4) antibody at 1:500 dilution and pH6 (Abcam plc). An antigen-retrieval step was performed using citrate buffer and a steamer for 30 min. For a negative control normal mouse serum was substituted for the antibody. Immunohistology sections were independently evaluated by 2 observers (LR, TG). Five randomly chosen fields of view were light-microscopically assessed at 250–400-fold magnification. Where disagreement occurred, decision by consensus was made. The immunoreactivity in the dermis was semi-quantitatively graded, as follows: 0 = none (0%), 1=slight (>0 ≤25%), 2 = moderate (>25 <75%), and 3 = strong (≥75–100%).
Data analysis was performed using the statistical package MedCalc Software (Mariakerke, Belgium). Data distribution was assessed by means of the D’Agostino-Pearson test. Based on distribution, data were expressed as means ± SD. The results were analysed using one-way analysis of variance including the Student-Newman-Keuls post-hoc test. Correlation analysis was performed using the Pearson procedure. Differences were considered significant when p < 0.05.
RESULTS
Fig. 1 shows examples of fibulin-4 and -5 staining in AN and MDE lesions as compared to healthy control (HC) skin. In HC dermis both proteins were prominently expressed in elastic tissue and blood vessels (Fig. 1b and d). In contrast, strongly reduced expression of fibulin-4 was observed in AN (Fig. 1a) and MDE (not shown), together with almost absent fibulin-5 expression in MDE (Fig. 1c). A semi-quantitative analysis of the immunostainings (score range: 0–3; Fig. S11) showed that the dermal expression of fibulin-4 was significantly (p = 0.001) decreased in MDE (1 ± 0.58 and AN (0.83 ± 0.39) compared with HC (1.87 ± 0.57). Moreover, dermal fibulin-5 expression in MDE (0.09 ± 0.24) was significantly diminished (p = 0.015) compared with both AN (0.43 ± 0.24) and HC (0.38 ± 0.27).
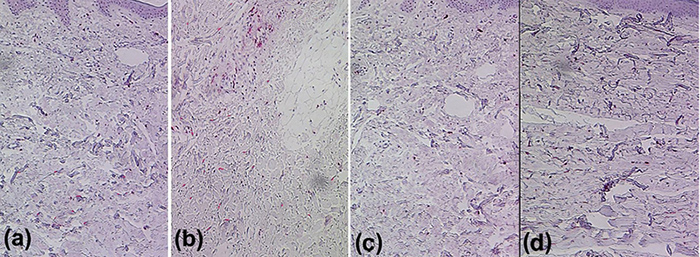
Fig. 1. (a) Strongly reduced dermal fibulin-4 in lesional skin of a patient with anetoderma, (c) absent dermal fibulin-5 immunoreactivity (red) in lesional skin of a patient with mid-dermal elastolysis, compared with (b, d) healthy controls. Original magnification ×250.
DISCUSSION
Previously, MMP9 has been linked to elastic tissue degradation in AN (2, 3), and Patroi et al. (4) observed intense intracytoplasmatic positivity for MMP9 protein expression in dendritic fibroblasts in the mid-dermis of patients with MDE. Furthermore, we have shown that MMP1 expression is also substantially increased in MDE (5–7). Thus, increased elastolytic activity by MMPs appears to represent an important pathogenic factor that might explain the loss of elastic fibres in MDE and AN (1–3). However, a significant impairment in re-synthesis and assembly of elastic fibres may also explain the permanent loss of elastic fibres in MDE and AN.
The genes for elastin and fibrillin-1, which are major components of elastin-associated microfibrils, remain active throughout life, but at the same time, elastogenesis in adults becomes relatively low (9, 10). The levels of LOX/LOXL mRNAs also decrease during adulthood (9–15). We have reported previously that affected elastin renewal due to altered LOXL2 expression may contribute to the pathogenesis of MDE (7).
Fibulin-4 and fibulin-5 belong to the most critical molecules for aiding assembly of elastic fibres. Indeed, recombinant fibulin-5 proteins accelerate elastic fibre assembly by dermal fibroblasts without changing the expression of genes related to the assembly, such as the soluble tropoelastin, LOX, fibrillin-1, and fibrillin-2 genes (9, 10). Moreover, adenovirus-mediated gene transfer of fibulin-5 gene is sufficient to regenerate elastic fibres in the mouse skin deficient in the fibulin-5 gene (15). Fibulin-4 deficiency in mice also leads to decreased tropoelastin expression; and a decrease in fibulin-4 expression may cause down-regulation of tropoelastin gene expression and impaired elastic-fibre formation (9–15).
We had the opportunity to study a relatively large sample size of MDE and AN and found that in both, dermal fibulin-4 protein expression was significantly decreased compared with controls. Moreover, dermal fibulin-5 was significantly diminished in MDE compared with controls and with AN. The results lend support to previous data indicating that not only an elastolytic overactivity, but also altered re-assembly of elastic fibres, probably play a significant role in the pathogenesis of MDE and AN. Limitations of the present study include the absence of gene expression analyses and functional studies.
ACKNOWLEDGEMENT
This study was supported by a FoRUM research grant of the Ruhr-University Bochum (FoRUM AZ: F715R-2011).
REFERENCES
1http://www.medicaljournals.se/acta/content/?doi=10.2340/00015555-2340


