


1Department of Dermatology, 2Department of Longevity Oral Science, Kagoshima University Graduate School of Medical and Dental Sciences, Kagoshima, and 3Department of Oral Disease Research, National Center for Geriatrics and Gerontology, Obu, Japan
Atopic dermatitis (AD) is a chronic inflammatory skin disease. Corticosteroids, which are widely used for AD treatment, have adverse effects, and alternative treatments are urgently needed. This study examined the effect of topical application of high-dose glucose on inflamed skin in a murine model of AD. High-dose glucose treatment on the ear reduced dermatitis scores and ear thicknesses in mite antigen-treated NC/Nga mice. The levels of thymus and activation-regulated chemokine (TARC), Th cytokines (interleukin (IL)-4, IL-5, IL-12, IL-13, and (interferon) IFN-γ), and IgE were decreased in the serum of high-dose glucose-treated mice. Expression of claudin-1 and filaggrin was reduced in the ear epithelium in the NC/Nga mice. How-ever, the reduced expression was restored by topical treatment with high-dose glucose. High-dose glucose also induced the expression of claudin-1 and filaggrin in cultured human skin keratinocytes. Co-stimulation with IL-4, IL-13, and thymic stromal lymphoprotein downregulated the expression of filaggrin in culture. However, high-dose glucose treatment restored the reduced expression of filaggrin. These results suggest that high-dose glucose treatment suppresses inflammation in the skin lesions by improving the skin barrier function.
Key words: claudin-1; filaggrin; sugar; steroid; anti-inflammation; allergy reaction; cytokines; IgE; skin barrier; mouse model.
Accepted Oct 2, 2017; Epub ahead of print Oct 2, 2017
Acta Derm Venereol 2018; 98: XX–XX.
Corr: Kenji Matsuhista, Department of Oral Disease Research, National Center of Geriatrics and Gerontology, 7-430 Morioka-machi, Obu, Aichi 747-8511, Japan. E-mail: kmatsu30@ncgg.go.jp; Takuro Kanekura, Department of Dermatology, Kagoshima University Graduate School of Medical and Dental Sciences, 8-35-1 Sakuragaoka, Kagoshima 890-8520, Japan. E-mail: takurok@m2.kufm.kagoshima-u.ac.jp
Atopic dermatitis (AD) is a chronic inflammatory skin disease characterized by increased IgE in serum, pruritic and relapsing eczematous skin lesions with a thickened epidermis, skin barrier defects, and infiltration of inflammatory cells, such as lymphocytes, eosinophils and mast cells (1–3). Thymus and activation-regulated chemokine (TARC), interleukin (IL)-4, IL-5, and IL-13 produced by Th2 cells are strongly associated with the onset and development of AD (4, 5). Th2 cells are associated with the acute phase of AD. On the other hand, Th1 cells, which produce IL-12 and interferon (IFN)-γ, contribute to the pathogenesis of the chronic phase of AD (6, 7). Corticosteroids have frequently been used for AD treatment (8). However, steroids have adverse effects in patients with AD (9) and alternative treatments are therefore needed.
Glucose is a major source of energy for mammalian cells. Abnormally high levels of glucose in the blood, as observed in diabetes, are harmful to the body and lead to the development of chronic complications, such as prolonged inflammation and impaired wound healing (10–13). However, topical application of sugars, such as honey and glycosaminoglycans, have wound-healing properties (14, 15). Glucose, one of the main sugars in honey, promotes wound healing (16–19). However, little is known about the effect of topical application of glucose on skin inflammation.
Recent reports suggest that declining epidermal barrier function influences the pathological characteristics of AD. Filaggrin is expressed in the granular layers as a > 400 kDa precursor protein, profilaggrin. Profilaggrin is cleaved into filaggrin monomers and N- and C-terminal peptides at the boundary of the stratum corneum and stratum granulosum. Filaggrin monomers are further degraded into natural moisturizing factors that are thought to maintain hydration of the upper stratum corneum and reduce the pH of the skin surface (20, 21). Filaggrin with both null mutations and loss-of-function variations is significantly associated with the development of AD in the European population (22). Tight junctions are also key contributors to the epidermal barrier, and claudin-1, which is a main component of tight junctions in the epidermis, is crucial for transepidermal water loss (TEWL) and skin barrier function. Claudin-1 is decreased in skin lesions of patients with AD, triggering skin inflammation (23).
To determine how glucose improves skin inflammation, we examined the effect of topical application of high-dose glucose on the inflamed skin in a murine model of chronic AD. We focused on claudin-1 and filaggrin and examined the expression of these molecules after treatment with a high concentration of glucose in vitro and in vivo.
NC/Nga mice (8-week-old males) bred under specific pathogen-free (SPF) conditions were purchased from Japan SLC Inc. (Shizuoka, Japan). The mice were given γ-ray-irradiated food (CRF-1, Oriental Bio Co., Kyoto, Japan) and distilled water (Otsuka Pharmaceutical Co., Ltd., Tokyo, Japan). This study was performed according to the Guidelines for Proper Conduct of Animal Experiments (Science Council in Japan, June 1, 2006), and the protocol was approved by the Institutional Animal Care Committee of KAC Co., Ltd. Throughout the experimental procedures, every effort was made to minimize the number of animals used and their suffering.
D-(+)-Glucose (G8270) was purchased from Sigma-Aldrich (St Louis, MO, USA). Prednisolone ointment (3 mg/g) was purchased from Kowa Co., Ltd (Nagoya, Japan). Tacrolimus 0.1% ointment was purchased from Astellas Pharma Inc. (Tokyo, Japan). White petrolatum was purchased from Maruishi Co., Ltd (Osaka, Japan).
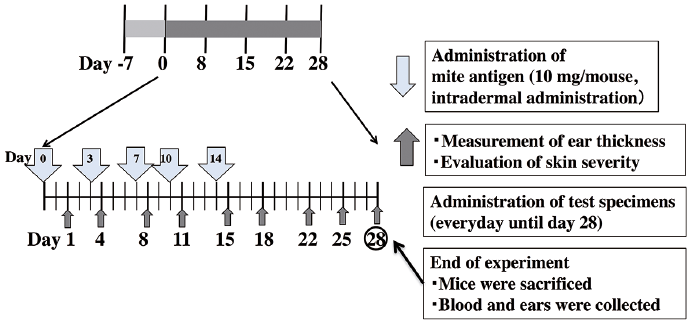
A dose of 100 mg mite cream (Biostir Inc., Kobe, Japan) impregnated with Dermatophagoides farina crude extract was injected intradermally to the right ear of each mouse twice a week, for 5 applications. From start the animals were divided into 5 groups with 6 mice in each group and topically treated with 20% D-glucose (50 mg/ear), prednisolone ointment (50 mg/ear), tacrolimus ointment (50 mg/ear), white petrolatum (50 mg/ear) or with phosphate-buffered saline (PBS) (mock group) every day for 4 weeks. The protocol is summarized in Fig. 1.
Fig. 1. Experimental design for the in vivo studies using the mouse model of atopic dermatitis.
Ear lesion severity was scored as described previously (24). Briefly, the clinical skin condition of the ears of the tested mice was examined macroscopically and scored every 3 or 4 days by 2 persons blinded to the treatment groups. A total clinical severity score for AD-like lesions was defined as the sum of the individual scores, graded as 0 (none), 1 (mild), 2 (moderate), and 3 (severe), for each of the 5 signs and symptoms (itch, erythema/haemorrhage, oedema, excoriation/erosion, and scaling/dryness). Ear thickness on the right ear of each mouse was measured 3 days a week using a thickness gauge (Mitsutoyo Corporation, Tokyo, Japan) (26). On day 28, the ears were photographed.
The ear skin was fixed with 10% buffered formalin solution (Wako Pure Chemical Industries, Ltd, Osaka, Japan), embedded in paraffin by the conventional method, and cut into 3–4 μm-thick sections. The sections were stained with haematoxylin and eosin (H&E) using Mayer’s Hematoxylin Solution. Mast cells and eosinophils in the sections were also stained with Toluidine Blue solution and Eosinophil-Mast Cell Stain Kit (ScyTek Laboratories, Inc., Logan, UT, USA), respectively.
The fixed ear skin was deparaffinized and hydrated in xylene and ethanol. After blocking with 10% normal goat serum for 30 min at room temperature, sections were incubated with avidin-conjugated anti-filaggrin antibody (1:500 diluent, Covance Inc., Princeton, NJ, USA) and avidin-conjugated anti-claudin-1 antibody (1:250 diluent, ab15098, Abcam, Cambridge, UK) overnight at 4°C. ABC Biotin solution (Vectastain ABC kit, Funakoshi Co., Ltd, Tokyo, Japan) and ABC reagent were subsequently used and the sections were then developed using DAB substrate.
Blood was collected on the last day of the experiment (day 28) and stored at −80°C until use. Serum IgE levels were measured by enzyme-linked immunoassay (ELISA) (Takara Bio Inc., Shiga, Japan) according to the manufacturer’s instructions. The levels of TARC, the Th2 cytokines, IL-4, IL-5, and IL-13, and the Th1 cytokines, IL-12 and IFN-γ, in sera were measured by ELISA (R&D Systems, Minneapolis, MN, USA).
Normal human skin keratinocytes (Lifeline Cell Technology, Frederick, MD, USA) were cultured in HuMeda-KG2 (Kurabo Industries, Osaka, Japan) at 37°C in a humidified atmosphere of 5% CO2. The cells were cultured until 70% confluence on 100-mm diameter culture dishes and then transferred to 6-well flat-bottom culture plates to perform the following experiments.
Confluent human skin keratinocytes in 6-well flat-bottom culture plates were incubated in HuMeda-KG2 overnight. The cells were then washed twice with PBS and incubated in HuMeda-KG2 supplemented with 6 or 110 mM D-glucose for 0–24 h. The cells were washed twice with PBS and then collected. In some experiments, confluent human skin keratinocytes in 6-well flat-bottom culture plates were incubated with 100 ng/ml IL-4 (R&D Systems, Minneapolis, MN, USA) and 100 ng/ml IL-13 (R&D Systems) in HuMeda-KG supplemented with 6 mM D-glucose for 72 h. The cells were then washed twice with PBS and incubated in HuMeda-KG2 supplemented with 110 mM D-glucose for 30–90 min. Confluent human skin keratinocytes were also incubated with 100 ng/ml thymic stromal lymphoprotein (TSLP) (Sigma) for 16 h and incubated in HuMeda-KG2 supplemented with 110 mM D-glucose for 30–90 min. The cells were washed twice with PBS and then collected.
Total RNA was extracted from cultured cells using Nucleo Spin RNA kit (Takara Bio Inc., Shiga, Japan) according to the manufacturer’s instructions. The purified RNA was converted to cDNA using a ReverTra Ace-α kit (Takara Bio Inc.) by a Takara PCR Thermal cycler. Real-time PCR was performed by using SYBR Green Real time PCR Master Mix-Plus (TOYOBO Co., Ltd, Tokyo, Japan) on a 7300 Real-time PCR system (Thermo Fisher Scientific, Waltham, MA, USA). The target-specific primers were designed by Primer-BLAST. The primer sequences were as follows: human filaggrin, 5’-TGA GGG CAC TGA AAG GCA AA-3’ (forward) and 5’-GCT GAG AGT GTC TAA ACC CG-3¢ (reverse); human claudin-1, 5’-TGC CCT ACT TTG CTG TTC CT-3’ (forward) and 5’-TGA TTT TCT CCT TTT GCC TCT GTG-3’ (reverse). The samples were normalized to the housekeeping gene, GAPDH, and the results were reported for each sample relative to the control. The primer’s sequences for the housekeeping gene, GAPDH, were: 5’-TTT GGT ATC GTG GAA GGA CTC A-3’ (forward) and 5’-CAG TAG AGG CAG GGA TGA TGT-3’ (reverse). These experiments were performed in triplicate for each condition. The value was presented as the fold change between the samples using the ΔΔCt method (25).
Cells were lysed with CelLytic M (Sigma) supplemented with a protease inhibitor (Merck Millipore, Darmstadt, Germany) and a phosphate inhibitor (Nacalai Tesque, Kyoto, Japan). The protein concentration was determined by a BCA protein assay kit (Thermo Fisher Scientific). Equal amounts of proteins were separated by a gradient gel (ATTO Co., Tokyo, Japan) and transferred to Immobilon-P transfer membranes. The membranes were blocked by 5% skim milk in Tween 20 (TBST) for 1 h at room temperature and incubated with primary antibodies overnight at 4°C with shaking in 2% skim milk in TBST and then with the secondary antibody anti-rabbit IgG-HRP (IBL, Gunma, Japan) or anti-mouse IgG-HRP (IBL). The primary antibodies used were: filaggrin (ab245884; Abcam), claudin-1 (ab15098; Abcam), signal transducer and activator of transcription 3 (STAT3) (4904P; Cell Signaling Technology, Inc., Danvers, MA, USA), p-STAT3 (9134P; Cell Signaling Technology) and β-actin (Loading control: GTX109639, Funakoshi). Immunoreactivity was detected using the ImmunoStar LD (Wako). Band intensity was measured using the CS Analyzer (ATTO) by the zone densitometry method. β-actin was used as a loading control.
Results are expressed as means ± standard deviations (SD). Significance between mean values was determined by one-way analysis of variance (ANOVA), followed by Tukey’s test for group comparisons, with p < 0.05 considered significant.
Topical application of a high concentration of glucose suppresses skin inflammation induced by mite antigen in NC/Nga mice
NC/Nga mice, in which AD was induced by topical application of mite antigen on the ears showed that the total clinical severity scores increased in all treatment-groups until Day 15. After mite antigen treatment was stopped, the severity scores significantly decreased in the glucose, prednisolone, and tacrolimus groups compared with those in the mock and white petrolatum groups (Fig. 2A). Similarly, ear thickness was significantly reduced by treatment with prednisolone ointment, high-dose glucose, and tacrolimus ointment (Fig. 2B). These results suggest that topical treatment with high-dose glucose is almost as effective as tacrolimus in allevating the AD-like lesions on the ears. Incidentally, erosion and bleeding were observed in the auricles of the mock group at Day 28, whereas the 20% D-glucose group only exhibited slight dilation of capillaries and redness of the skin (Fig. S1).
Fig. 2. Topical application of a high concentration of glucose alleviates atopic dermatitis symptoms in mite antigen-sensitized NC/Nga mice. (A) Clinical score and (B) thickness of the mite antigen-treated right ear of each NC/Nga mouse were determined. The clinical skin score was calculated as the sum of scores for 5 clinical symptoms. The ear thickness was measured by a dial thickness gauge. All values are expressed as mean ± SD of 6 mice per group. *,**p < 0.05, p < 0.01 compared with the mock group. Mock: stimulation with mite antigen and treatment with phosphate-buffered saline.
To assess the histopathology of the skin, ears obtained at Day 28 from glucose-, prednisolone-, mock-treated and negative control mice were sectioned and stained with H&E or specific stainings for mast cells and eosinophils (Fig. S2). Skin lesions of mite-treated NC/Nga mice presented hyperkeratosis, acanthosis, erosion, and infiltration of inflammatory cells. However, the number of infiltrating cells in the dermis decreased significantly in response to high-dose glucose and prednisolone treat-ments. Increase of mast cells was observed in the auricular skin due to the application of mite antigen, but the level tended to decrease after application of high concentration of D-glucose. Similarly, infiltration of eosinophils was remarkably attenuated by treatment with prednisolone and high concentration D-glucose. These results suggest that treatment with high concentration D-glucose attenuates the infiltration of inflammatory cells, a characteristic of AD.
Since the inflammation process involves different cytokines (26) and various cytokines produced by Th1 and Th2 cells contribute to the onset and progression of AD, we examined the serum levels of several chemokines and cytokines in mite antigen-stimulated NC/Nga mice. The level of chemokine TARC was markedly increased at Day 28 in mock-treated animals but was reduced by treatment with 20% D-glucose or prednisolone (Fig. 3A). Similarly, the increased level of IgE in the serum of mite antigen-stimulated NC/Nga mice was reduced by high-dose glucose and prednisolone treatment (Fig. 3B). Furthermore, the levels of Th1 cytokines, IL12, IFN-γ, and TNF-α, and Th2 cytokines, IL-4, IL-5, IL-6 and IL-13, which were significantly increased in the serum of mite antigen-treated NC/Nga mice, were also reduced by 20% D-glucose and prednisolone treatment (Fig. 4). These results suggest that topical treatment with high-dose glucose suppresses the production of cytokines by T cells and infiltrating immune cells, reducing the skin inflammation in the AD model.
Fig. 3. High-dose glucose treatment suppresses (A) mite antigen-induced Thymus and Activation-Regulated Chemokine (TARC) production and (B) IgE production in NC/Nga mice. Blood from mite antigen-stimulated NC/Nga mice after treatment with prednisolone 20% D-glucose, or a vehicle (Mock) and from non-stimulated NC/Nga mice (Control) was collected on the last day of the experiment (day 28) and serum levels of TARC and IgE were measured by enzyme-linked immunoassay (ELISA). n = 6 ± standard deviation (SD), **p < 0.01; *p < 0.05.
Fig. 4. High-dose glucose treatment suppresses mite-induced production of Th1 and Th2 cytokines in NC/Nga mice. Blood from mite antigen-stimulated NC/Nga mice after treatment with prednisolone 20% D-glucose, and a vehicle (Control) was collected on the last day of the experiment (day 28) and levels of serum cytokines (interleukin (IL)-4, IL-5, IL-6, IL-12, IL-13, interferon (IFN)-γ and tumour necrosis factor alpha (TNF-α)) were measured by enzyme-linked immunoassay (ELISA). n = 6 ± standard deviation (SD), **p < 0.01; *p < 0.05.
We next explored the mechanism by which high-dose glucose treatment improves skin inflammation, taking into account that epidermal barrier dysfunction and immune abnormalities are observed in patients with AD (27, 28). We first examined the epidermal expression of claudin-1 and filaggrin, which are associated with skin barrier function. Indeed, the expression of claudin-1 in the granular layer was decreased and progressively disappeared in the mite antigen-exposed mice (Fig. S3). Treatment with prednisolone rescued claudin-1 expression, but it was now located in the nuclei. In contrast, a strong and continuous expression of claudin-1 was observed in the ear sections of the 20% D-glucose group. Epidermal expression of filaggrin was reduced in the ears from mite antigen-treated mice. However, the expression was broader and stronger in the granular layer of the ears from the 20% D-glucose group. These results suggest that reduced expression of claudin-1 and filaggrin in the AD model was recovered by treatment with a high concentration of glucose.
We next examined whether a high concentration of glucose induces claudin-1 and filaggrin expression also in cultured skin keratinocytes. Human skin keratinocytes were incubated with a high concentration of D-glucose and collected to measure the levels of claudin-1 and filaggrin. A high concentration of glucose induced mRNA expression of claudin-1 and filaggrin in the keratinocytes (Fig. 5A). The claudin-1 expression peaked after 0.5-h treatment and filaggrin expression peaked 2 h after addition of 110 mM D-glucose. We also examined the effects of various concentrations (6, 55, 110, and 165 mM) of glucose on the expression of claudin-1 and filaggrin by western blot analysis (Fig. 5B). The expression of claudin-1 was strongly induced by 55 mM D-glucose, while filaggrin expression was induced by 165 mM D-glucose. In a time course study, increased expression of claudin-1 and filaggrin in the culture was observed 2 h after the addition of a high concentration of D-glucose and the high expression levels persisted until 24 h (Fig. 5B).
Fig. 5. High-dose glucose treatment induces the expression of filaggrin and claudin-1 in human skin keratinocytes in vitro. (A) Human skin keratinocytes were treated with 110 mM D-glucose for 0, 0.5, 1 2, 3, 4 and 5 h, and total RNA was extracted. The levels of filaggrin and claudin-1 mRNA expression were measured by quantitative real-time reverse transcription PCR. n=3 ± standard deviation (SD) (B) Human keratinocytes were treated with 165 mM (for filaggrin) and 55 mM (for claudin-1) of D-glucose for 0 2, 4, 6, and 24 h. Total lysates were immunoblotted with antibodies against filaggrin, claudin-1 and β-actin.
We then examined the effects of TSLP, IL-4, and IL-13, i.e. cytokines involved in AD-associated inflammation, on the expression of claudin-1 and filaggrin. Filaggrin expression was decreased by TSLP stimulation, but the reduced expression was reverted by addition of 110 mM D-glucose for 30–90 min after stimulation (Fig. 6A). In addition, STAT3 phosphorylation was increased in human skin keratinocytes stimulated with TSLP, and the level was reduced by treatment with 110 mM of D-glucose (Fig. 6A). Co-stimulation with IL-4 and IL-13 reduced the expression of filaggrin in cells, but the expression was augmented by treatment with 55 mM D-glucose for 90 min after IL-4 and IL-13 stimulation (Fig. 6B). These results suggest that a high concentration of D-glucose can restore the expression of barrier-related proteins, such as claudin-1 and filaggrin.
Fig. 6. High-dose glucose treatment restores reduced production of filaggrin in human skin keratinocytes in vitro. (A) Human skin keratinocytes were stimulated with 100 ng/ml of thymic stromal lymphoprotein (TSLP) for 16 h and then treated with 110 mM D-glucose for 30–90 min. Total cell lysates were immunoblotted with antibodies against filaggrin, phospho-STAT3, total STAT3, and β-actin. (B) Human skin keratinocytes were stimulated with 100 ng/ml interleukin (IL)-4 and 100 ng/ml IL-13 for 72 h and were then treated with 110 mM D-glucose for 30–90 min. Total cell lysates were immunoblotted with an antibody against filaggrin and β-actin.
The toxicity of high levels of glucose in the blood is well known (29, 30). However, little is known about the effect of topical application of a high concentration of glucose. This study demonstrated that topical application of high-dose glucose improves AD-like skin lesions induced by mite antigen and exhibits potent anti-inflammatory activities. It also showed that high-dose glucose induced the expression of skin barrier-associated proteins, such as claudin-1 and filaggrin, in AD-like skin lesions of mice and in cultured human skin keratinocytes. Furthermore, the reduction in filaggrin expression induced by cytokines was restored by treatment with a high concentration of glucose in vitro. These results suggest that topical application of a high concentration of glucose may improve AD-like lesions by inducing filaggrin and claudin-1 expression, supporting the recovery of skin barrier function.
Our overall results indicate that topical application of a high concentration of glucose is associated with improvement in inflammation and skin barrier function. However, the mechanism by which a high concentration of glucose improves skin inflammation is unclear, as is its molecular mechanism in reducing the production of cytokines and IgE in mite antigen-stimulated mice. TARC is a chemokine involved in Th2 cell activation and migration into the skin. TARC level in serum is considered to be an important indicator of AD activity (31). In addition, IL-4, IL-5, and IL-13 produced by Th2 cells induce immune responses in the early phase of AD (4, 5). Furthermore, these Th2 cytokines, especially IL-4, are essential for the production of IgE. These allergen/IgE/Th2 cytokines cooperatively enhance skin inflammation in AD and were reduced by topically applied high-dose glucose in AD model. Similarly, treatment with a high concentration of glucose suppressed the production of Th1 cytokines, including IL-12 and IFN-γ, which are thought to inhibit the function of Th2 cells, followed by reduction in allergic inflammation in AD. In contrast, some researchers showed that Th1 cells contribute to the pathogenesis of the chronic phase of AD (6, 7). There-fore, suppression of the production of Th1 cytokines by a high concentration of glucose may exacerbate skin inflammation. We have not examined whether a high concentration of glucose directly inhibits the production of these Th cytokines. Th1 cells, Th2 cells, and Th17 cells express high levels of the glucose transporter, Glut1, and present a high glycolytic metabolism (32, 33). Therefore, high levels of glucose uptake and glycolysis in Th cells may induce cell differentiation and cytokine production. The level of claudin-1, a key molecule for epithelial tight junctions, is reduced in the skin of AD (34). Mutations in the filaggrin gene result in impaired skin barrier function and are strongly associated with the development of AD (35). In addition, immune-mediated skin inflammation is also observed in AD patients with acquired filaggrin decrease (36). Following loss of filaggrin expression, severe AD pathology and increased TEWL occurs. In addition, changes in the expression of various genes, including inflammation-related genes, occur in relation to the extent of filaggrin mRNA expression (37). High levels of Th1 and Th2 cytokines were observed in the serum of AD patients with reduced expression levels of claudin-1 and filaggrin in skin lesions (38). These skin barrier defects would sustain allergen entry, leading to Th cell infiltration, and may cause inflammation in AD. Therefore, recovery of the skin barrier function by topical treatment with a high concentration of glucose may reduce the production of Th cytokines and IgE.
Keratins are structural proteins involved in the formation of the epithelial cytoskeleton and stabilize epithelial cells (39). A decrease in the expression of keratins may reduce epithelial integrity. Restorative expression of keratin 5, which is a marker of mitotically active basal cells in the epidermis, was observed in mice treated with a high concentration of glucose (Fig. S4).
These results indicate that epithelial stabilization induced by a high concentration of glucose may contribute to restoring the skin barrier function in mite antigen-stimulated mice and to the suppression of mite antigen-induced inflammatory responses.
While the results of the current study indicate that a high concentration of glucose induces the expression of claudin-1 and filaggrin, the underlying mechanism in this study could not be clarified. Glucose transporter 1 (Glut1) is involved in the formation of tissue barrier of the endothelium and epithelium. Glut1 is a maturation marker of the blood-brain barrier (BBB) and is involved in the formation of tight junctions (40). Glut1 is expressed in the skin epithelium and is excessively expressed in psoriatic skin (41). Therefore, a high concentration of glucose is efficiently taken up into cells in inflamed skin lesions, and the expression of tight junction-related molecules, including claudin-1, may then be induced. While topical application of a high concentration of glucose restored filaggrin expression reduced by TSLP stimulation, the underlying mechanism is unknown. TSLP is an IL-7-like cytokine that activates dendritic cells to induce a Th2 response (42). TSLP is highly expressed in acute and chronic AD lesions and is associated with the development of AD. TSLP decreases the expression of filaggrin by activating STAT3 and ERK phosphorylation (43). Therefore, treatment with a high concentration of glucose may suppress the reduction of filaggrin expression by TSLP through restoration of STAT3 activation. Recent reports have indicated that the transcription factor, MafB, regulates the expression of filaggrin in human keratinocytes. Expression of the filaggrin gene was reduced in the epidermis of MafB-deficient mice. Furthermore, the expression of filaggrin and MafB was significantly decreased in patients with AD and psoriasis (44). MafB induces filaggrin gene expression by binding to MARE-like-1 element within the ECR2 promoter region. MafB is a crucial factor for glucose sensing and insulin secretion in islet β cells (45). Therefore, a high concentration of glucose may promote gene expression of filaggrin by activating MafB in epithelial keratinocytes.
In conclusion, this study revealed that topical treatment with high-dose glucose ameliorates AD inflammation in a murine model. Topical steroids, such as prednisolone, and immunomodulators, such as FK506, are widely used for AD treatment; however, these drugs have adverse effects (9). A safe and effective treatment for AD is there-fore needed. Our observations may lead to a promising new treatment for patients with AD.
This work was supported by a Grant-in-Aid for Scientific Research B (to K.M.) and a Grant-in-Aid for Challenging Exploratory Research (to K.M.) from the Ministry of Education, Culture, Sports, Science and Technology, Japan.


 Click to show fullsize
Click to show fullsize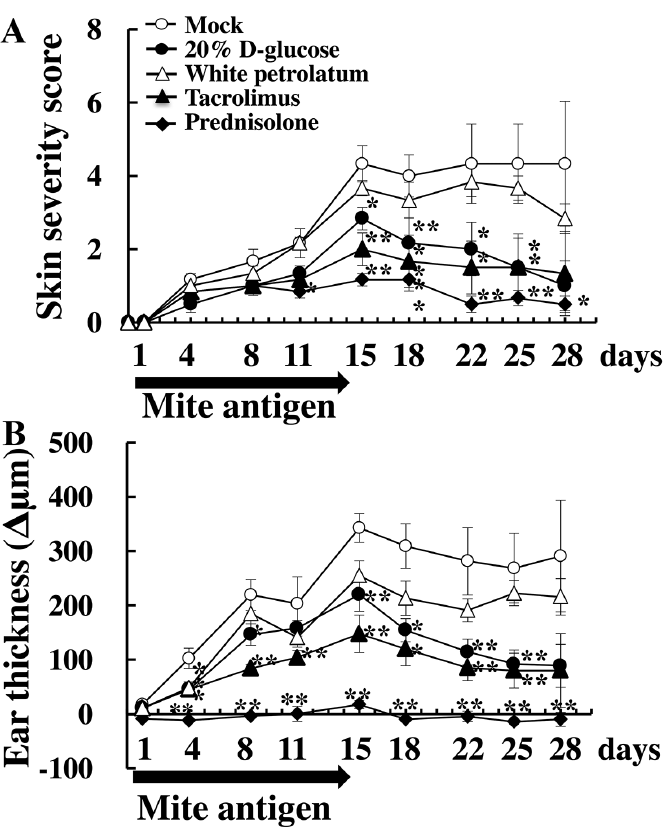 Click to show fullsize
Click to show fullsize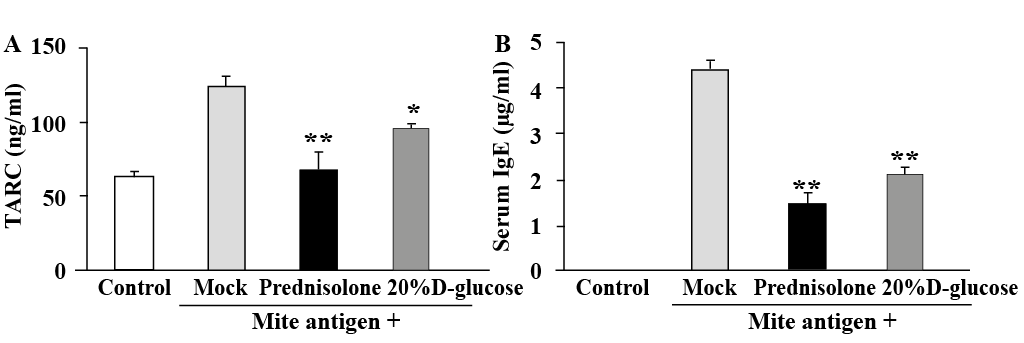 Click to show fullsize
Click to show fullsize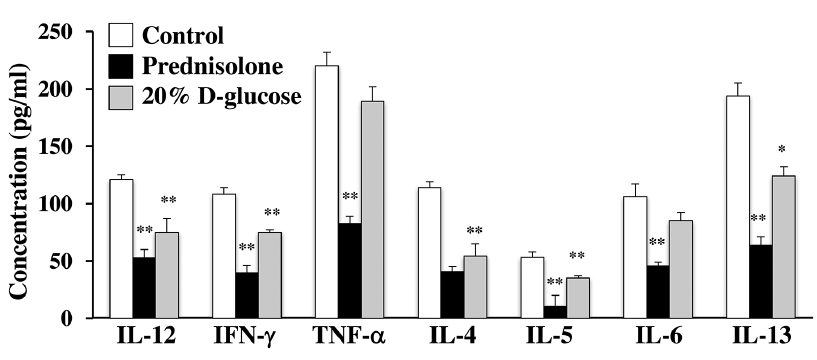 Click to show fullsize
Click to show fullsize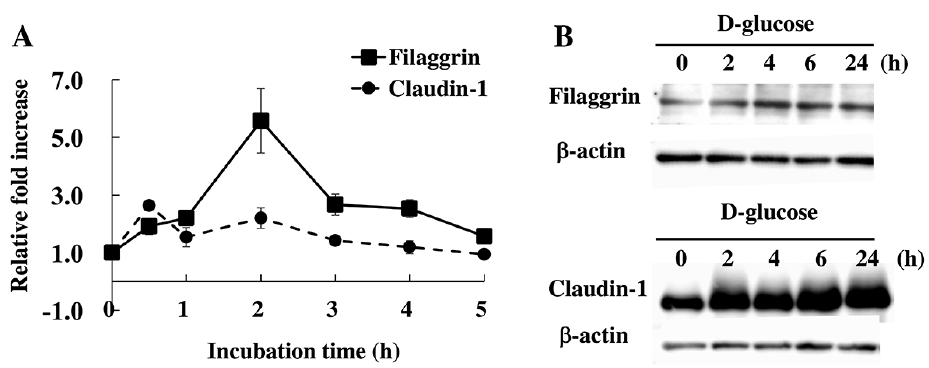 Click to show fullsize
Click to show fullsize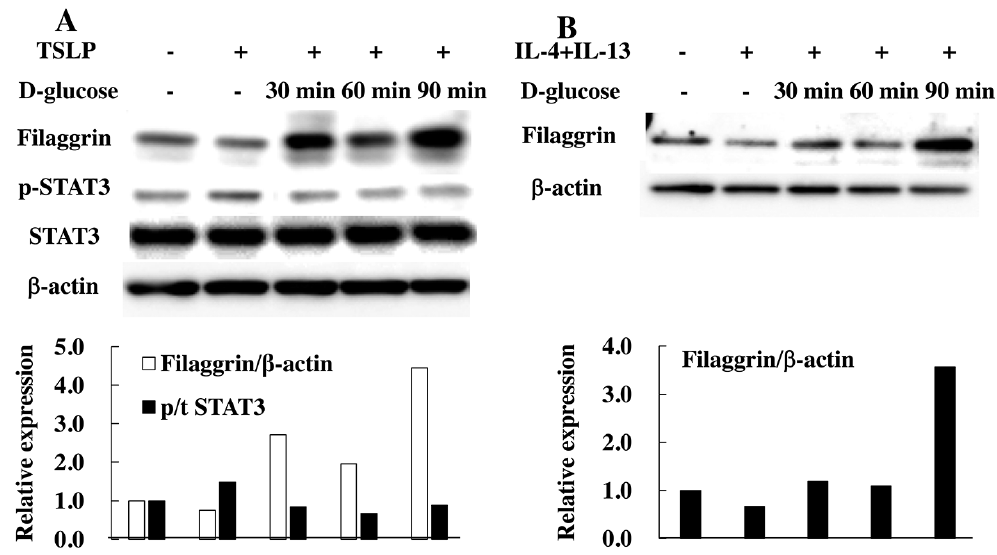 Click to show fullsize
Click to show fullsize